15.1Metabolism Is Composed of Many Coupled, Interconnecting Reactions
Metabolism Is Composed of Many Coupled, Interconnecting Reactions
Living organisms require a continual input of free energy for three major purposes: (1) the performance of mechanical work in muscle contraction and cellular movements, (2) the active transport of molecules and ions, and (3) the synthesis of macromolecules and other biomolecules from simple precursors. The free energy used in these processes, which maintain an organism in a state that is far from equilibrium, is derived from the environment. Photosynthetic organisms, or phototrophs, obtain this energy by trapping sunlight, whereas chemotrophs, which include animals, obtain energy through the oxidation of foodstuffs generated by phototrophs.
Metabolism consists of energy-yielding and energy-requiring reactions
Metabolism is essentially a sequence of chemical reactions that begins with a particular molecule and results in the formation of some other molecule or molecules in a carefully defined fashion (Figure 15.1). There are many such defined pathways in the cell (Figure 15.2), and we will examine a few of them in some detail later. These pathways are interdependent, and their activity is coordinated by exquisitely sensitive means of communication in which allosteric enzymes are predominant (Section 10.1). We considered the principles of this communication in Chapter 14.
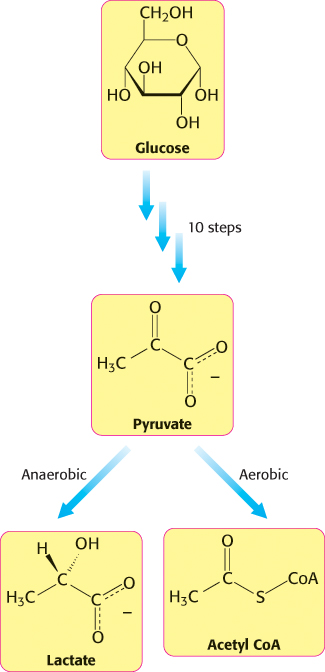
We can divide metabolic pathways into two broad classes: (1) those that convert energy from fuels into biologically useful forms, and (2) those that require inputs of energy to proceed. Although this division is often imprecise, it is nonetheless a useful distinction in an examination of metabolism. Those reactions that transform fuels into cellular energy are called catabolic reactions or, more generally, catabolism.
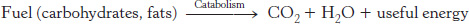
Those reactions that require energy—
425
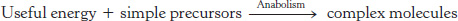
Some pathways can be either anabolic or catabolic, depending on the energy conditions in the cell. These pathways are referred to as amphibolic pathways.
An important general principle of metabolism is that biosynthetic and degradative pathways are almost always distinct. This separation is necessary for energetic reasons, as will be evident in subsequent chapters. It also facilitates the control of metabolism.
A thermodynamically unfavorable reaction can be driven by a favorable reaction
How are specific pathways constructed from individual reactions? A pathway must satisfy minimally two criteria: (1) the individual reactions must be specific, and (2) the entire set of reactions that constitute the pathway must be thermodynamically favored. A reaction that is specific will yield only one particular product or set of products from its reactants. As discussed in Chapter 8, enzymes provide this specificity. The thermodynamics of metabolism is most readily approached in relation to free energy, which was discussed in chapters 1 and 8. A reaction can occur spontaneously only if ΔG, the change in free energy, is negative. Recall that ΔG for the formation of products C and D from substrates A and B is given by
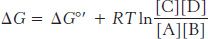
Thus, the ΔG of a reaction depends on the nature of the reactants and products (expressed by the ΔG°′ term, the standard free-
426
An important thermodynamic fact is that the overall free-
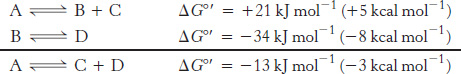
Under standard conditions, A cannot be spontaneously converted into B and C, because ΔG°′ is positive. However, the conversion of B into D under standard conditions is thermodynamically feasible. Because free-