22.6Acetyl CoA Carboxylase Plays a Key Role in Controlling Fatty Acid Metabolism
Acetyl CoA Carboxylase Plays a Key Role in Controlling Fatty Acid Metabolism
Fatty acid metabolism is stringently controlled so that synthesis and degradation are highly responsive to physiological needs. Fatty acid synthesis is maximal when carbohydrates and energy are plentiful and when fatty acids are scarce. Acetyl CoA carboxylase 1 and 2 play essential roles in regulating fatty acid synthesis and degradation. Recall that this enzyme catalyzes the committed step in fatty acid synthesis: the production of malonyl CoA (the activated two-
671
Acetyl CoA carboxylase is regulated by conditions in the cell
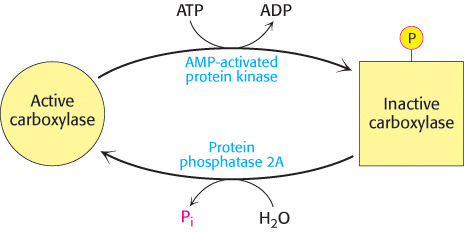
Acetyl CoA carboxylase responds to changes in its immediate environment. Acetyl CoA carboxylase is switched off by phosphorylation and activated by dephosphorylation (Figure 22.34). AMP-
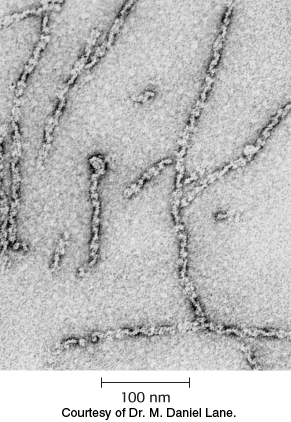
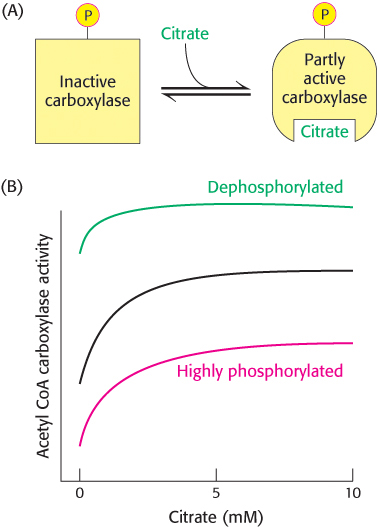
The carboxylase is also allosterically stimulated by citrate. The level of citrate is high when both acetyl CoA and ATP are abundant, signifying that raw materials and energy are available for fatty acid synthesis. Citrate acts in an unusual manner on inactive acetyl CoA carboxylase, which exists as isolated inactive dimers. Citrate facilitates the polymerization of the inactive dimers into active filaments (Figure 22.35). However, polymerization by citrate alone requires supraphysiological concentrations. In the cell, citrate-
The isozyme acetyl CoA carboxylase 2, located in the mitochondria, plays a role in the regulation of fatty acid degradation. Malonyl CoA, the product of the carboxylase reaction, is present at a high level when fuel molecules are abundant. Malonyl CoA inhibits carnitine acyltransferase I, preventing the entry of fatty acyl CoAs into the mitochondrial matrix in times of plenty. Malonyl CoA is an especially effective inhibitor of carnitine acyltransferase I in heart and muscle, tissues that have little fatty acid synthesis capacity of their own. In these tissues, acetyl CoA carboxylase may be a purely regulatory enzyme.
Acetyl CoA carboxylase is regulated by a variety of hormones
Acetyl CoA carboxylase is controlled by the hormones glucagon, epinephrine, and insulin, which denote the overall energy status of the organism. Insulin stimulates fatty acid synthesis by activating the carboxylase, whereas glucagon and epinephrine have the reverse effect.
Regulation by glucagon and epinephrine. Consider, again, a person who has just awakened from a night’s sleep and begins a bout of exercise. As mentioned, glycogen stores will be low, but lipids are readily available for mobilization.
As stated earlier, the hormones glucagon and epinephrine, present under conditions of fasting and exercise, will stimulate the release of fatty acids from triacylglycerols in fat cells, which will be released into the blood, and probably from muscle cells, where they will be used immediately as fuel. These same hormones will inhibit fatty acid synthesis by inhibiting acetyl CoA carboxylase. Although the exact mechanism by which these hormones exert their effects is not known, the net result is to augment the inhibition by the AMP-
672
Regulation by insulin. Now consider the situation after the exercise has ended and the runner has had a meal. In this case, the hormone insulin inhibits the mobilization of fatty acids and stimulates their accumulation as triacylglycerols by muscle and adipose tissue. Insulin also stimulates fatty acid synthesis by activating acetyl CoA carboxylase. Insulin activates the carboxylase by enhancing the phosphorylation of AMPK by protein kinase B, which inhibits AMPK, as well as by stimulating the activity of a protein phosphatase that dephosphorylates and activates acetyl CoA carboxylase. Thus, the signal molecules glucagon, epinephrine, and insulin act in concert on triacylglycerol metabolism and acetyl CoA carboxylase to carefully regulate the utilization and storage of fatty acids.
Response to diet. Long-