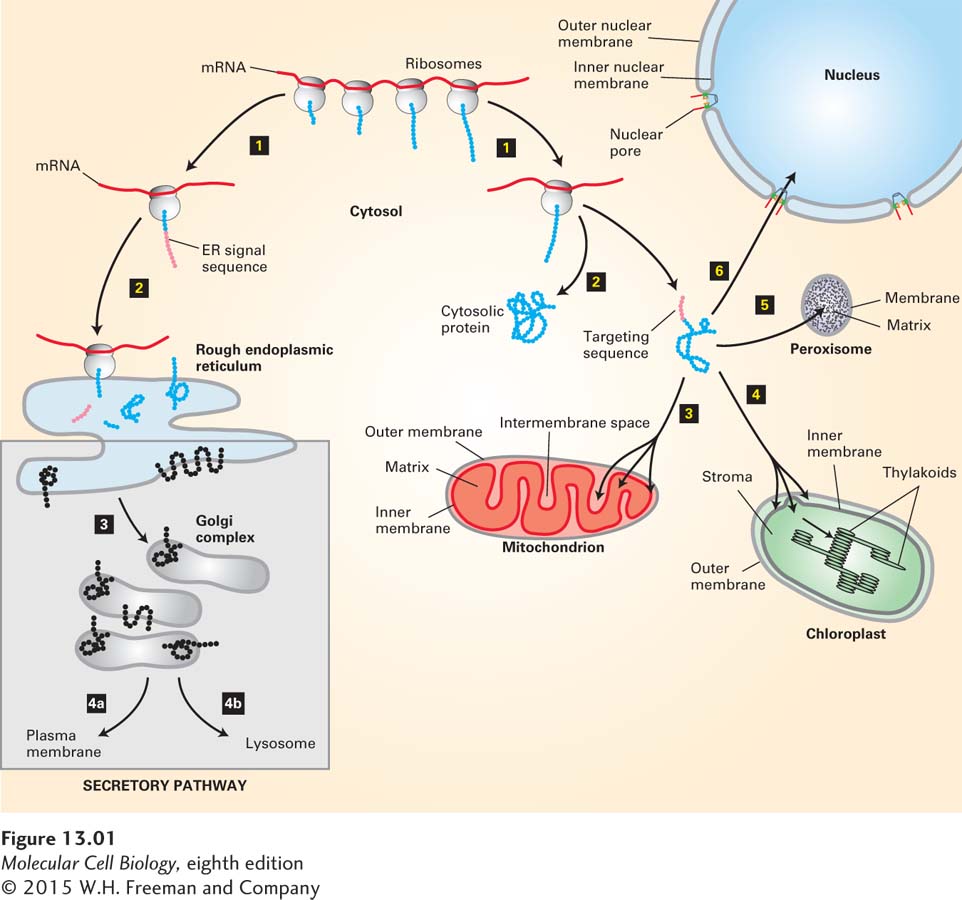
The delivery of newly synthesized proteins to their proper cellular destinations, usually referred to as protein targeting or protein sorting, encompasses two very different kinds of processes: signal-based targeting and vesicle-based trafficking. The first kind of process involves the targeting of a newly synthesized protein from the cytoplasm to an intracellular organelle. Targeting can occur during translation or soon after synthesis of the protein is complete. For membrane proteins, targeting leads to insertion of the protein into the lipid bilayer of the membrane, whereas for water-soluble proteins, targeting leads to translocation of the entire protein across the membrane into the aqueous interior of the organelle. Proteins are sorted to the endoplasmic reticulum (ER), mitochondria, chloroplasts, peroxisomes, and nucleus by this general process (Figure 13-1).
FIGURE 13-1Overview of major protein-sorting pathways in eukaryotes. All nuclear DNA–encoded mRNAs are translated on cytosolic ribosomes. Right (nonsecretory pathways): Synthesis of proteins lacking an ER signal sequence is completed on free ribosomes (step 1). Those proteins that contain no targeting sequence are released into the cytosol and remain there (step 2). Proteins with an organelle-specific targeting sequence (pink) are first released into the cytosol (step 2) but are then imported into mitochondria, chloroplasts, peroxisomes, or the nucleus (steps 3–6). Mitochondrial and chloroplast proteins typically pass through the outer and inner membranes to enter the matrix or stromal space, respectively. Other proteins are sorted to other subcompartments of these organelles by additional sorting steps. Nuclear proteins enter and exit through visible pores in the nuclear envelope. Left (secretory pathway): Ribosomes synthesizing nascent proteins in the secretory pathway are directed to the rough endoplasmic reticulum (ER) by an ER signal sequence (pink; steps 1 and 2). After translation is completed on the ER, these proteins can move via transport vesicles to the Golgi complex (step 3). Further sorting delivers proteins either to the plasma membrane or to lysosomes (step 4a or 4b). The vesicle-based processes underlying the secretory pathway (steps 3 and 4, shaded box) are discussed in Chapter 14.
The second general sorting process, known as the secretory pathway, involves transport of proteins from the ER to their final destination within membrane-enclosed vesicles. For many proteins, including those that make up the extracellular matrix, the final destination is the outside of the cell (hence the name); integral membrane proteins are also transported to the Golgi complex, lysosomes, and plasma membrane by this process. The secretory pathway begins in the ER; thus all proteins slated to enter the secretory pathway are initially targeted to this organelle.
Credits Always Shown
Part (a) courtesy of Dr. Marilyn G. Farquhar, University of California, San Diego.
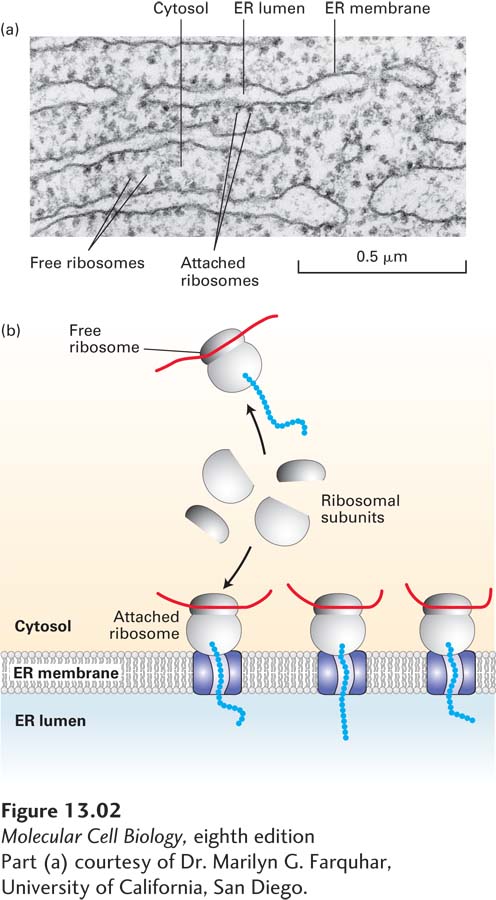
FIGURE 13-2Structure of the rough ER. (a) Electron micrograph of ribosomes attached to the rough ER in a pancreatic acinar cell. Most of the proteins synthesized by this type of cell are secretory proteins and are formed on membrane-attached ribosomes. A few unattached (free) ribosomes are evident; presumably, these ribosomes are synthesizing cytosolic or other nonsecretory proteins. (b) Schematic representation of protein synthesis on the ER. Note that membrane-bound and free cytosolic ribosomes are identical. Membrane-bound ribosomes are recruited to the ER during synthesis of a polypeptide containing an ER signal sequence.
Biochemical studies using a cell-free protein-synthesizing system, mRNA encoding a secretory protein, and microsomes stripped of their own bound ribosomes have elucidated the function and fate of ER signal sequences. Initial experiments with this system demonstrated that a typical secretory protein is incorporated into microsomes and has its signal sequence removed only if the microsomes are present during protein synthesis. If microsomes are added to the system after protein synthesis is completed, no protein transport into the microsomes occurs (Figure 13-4). Subsequent experiments were designed to determine the precise stage of protein synthesis at which microsomes must be present in order for translocation to occur. In these experiments, microsomes were added to the reaction mixtures at different times after protein synthesis had begun. These experiments showed that microsomes must be added before the first 70 or so amino acids are translated in order for the completed secretory protein to be localized in the microsomal lumen. At this point, the first 40 or so amino acids protrude from the ribosome, including the signal sequence that will later be cleaved off, and the next 30 or so amino acids are still buried within a channel in the ribosome (see Figure 5-26). Thus the transport of most secretory proteins into the ER lumen begins while the incompletely synthesized (nascent) protein is still bound to the ribosome, a process referred to as cotranslational translocation.
Biochemical studies using a cell-free protein-synthesizing system, mRNA encoding a secretory protein, and microsomes stripped of their own bound ribosomes have elucidated the function and fate of ER signal sequences. Initial experiments with this system demonstrated that a typical secretory protein is incorporated into microsomes and has its signal sequence removed only if the microsomes are present during protein synthesis. If microsomes are added to the system after protein synthesis is completed, no protein transport into the microsomes occurs (Figure 13-4). Subsequent experiments were designed to determine the precise stage of protein synthesis at which microsomes must be present in order for translocation to occur. In these experiments, microsomes were added to the reaction mixtures at different times after protein synthesis had begun. These experiments showed that microsomes must be added before the first 70 or so amino acids are translated in order for the completed secretory protein to be localized in the microsomal lumen. At this point, the first 40 or so amino acids protrude from the ribosome, including the signal sequence that will later be cleaved off, and the next 30 or so amino acids are still buried within a channel in the ribosome (see Figure 5-26). Thus the transport of most secretory proteins into the ER lumen begins while the incompletely synthesized (nascent) protein is still bound to the ribosome, a process referred to as cotranslational translocation.
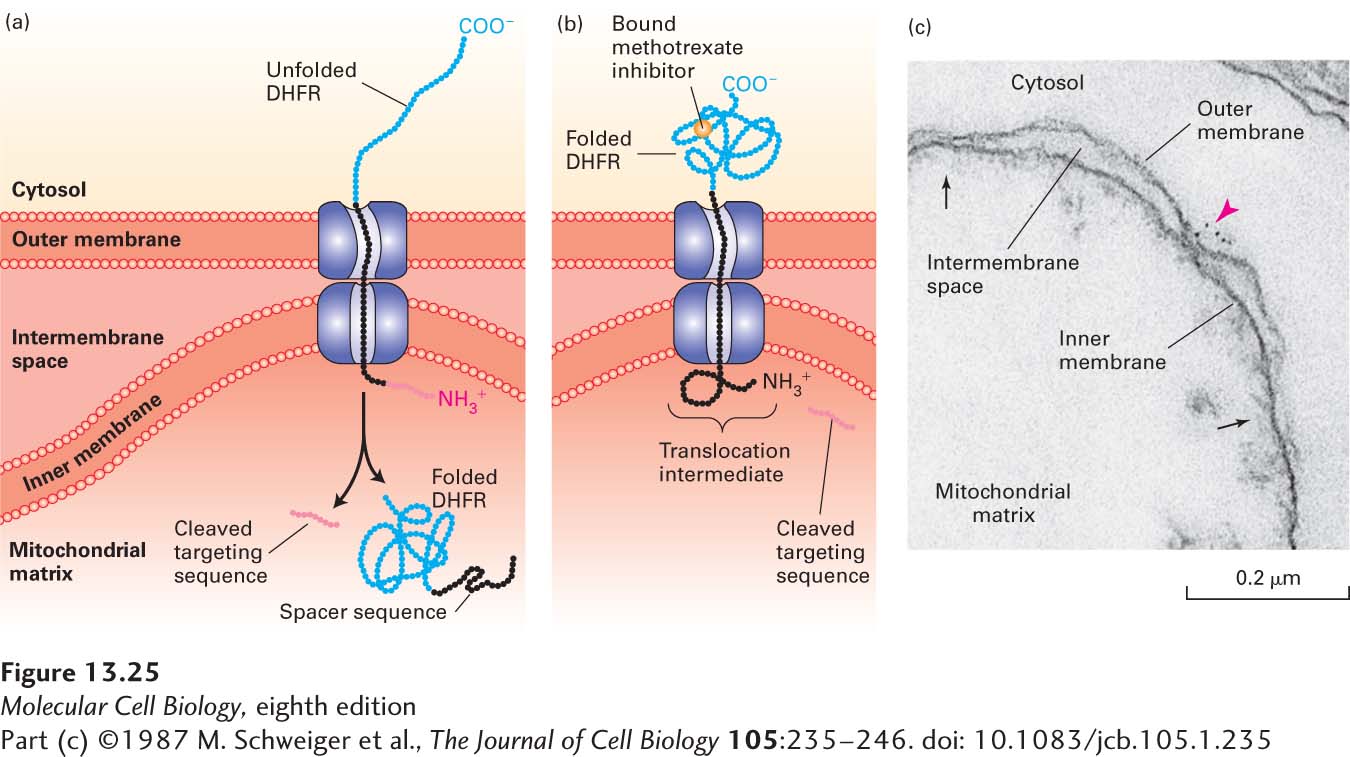
EXPERIMENTAL FIGURE 13-25Experiments with chimeric proteins elucidate mitochondrial protein import processes. These experiments show that a matrix-targeting sequence alone directs proteins to the mitochondrial matrix and that only unfolded proteins are translocated across both mitochondrial membranes. The chimeric protein in these experiments contained a matrix-targeting signal at its N-terminus (red), followed by a spacer sequence of no particular function (black), and then by dihydrofolate reductase (DHFR), an enzyme normally present only in the cytosol. (a) When the DHFR segment is unfolded, the chimeric protein moves across both membranes to the matrix of an energized mitochondrion, and the matrix-targeting signal is then removed. (b) When the C-terminus of the chimeric protein is locked in the folded state by binding of methotrexate, translocation is blocked. If the spacer sequence is long enough to extend across both transport channels, a stable translocation intermediate, with the targeting sequence cleaved off, is generated in the presence of methotrexate, as shown here. (c) The C-terminus of the translocation intermediate in (b) can be detected by incubating the mitochondria with antibodies that bind to the DHFR segment, followed by gold particles coated with bacterial protein A, which binds nonspecifically to antibody molecules (see Figure 4-33). An electron micrograph of a sectioned sample reveals gold particles (red arrowhead) bound to the translocation intermediate at a contact site between the inner and outer membranes. Other contact sites (black arrows) are also evident. See J. Rassow et al., 1990, FEBS Lett.275:190.