module 21 Community Succession
Even without human activity, natural communities do not stay the same forever. Change in the species composition of communities over time is a perpetual process in nature. In this module we will look at how both terrestrial and aquatic communities change over time.
Learning Objectives
After reading this module you should be able to
explain the process of primary succession.
explain the process of secondary succession.
explain the process of aquatic succession.
describe the factors that determine the species richness of a community.
Primary succession starts with no soil
Ecological succession The predictable replacement of one group of species by another group of species over time.
Virtually every community experiences ecological succession, which is the predictable replacement of one group of species by another group of species over time. Depending on the community type, ecological succession can occur over time spans varying from decades to centuries.
Primary succession Ecological succession occurring on surfaces that are initially devoid of soil.
Some terrestrial communities begin with bare rock and no soil. For example, a community may begin forming on newly exposed rock left behind after a glacial retreat, newly cooled lava after a volcanic eruption, or an abandoned parking lot. When succession begins with bare rock and no soil, we call it primary succession.
FIGURE 21.1 shows the process of primary succession in a temperate forest biome in New England. The bare rock can be colonized by organisms such as algae, lichens, and mosses—
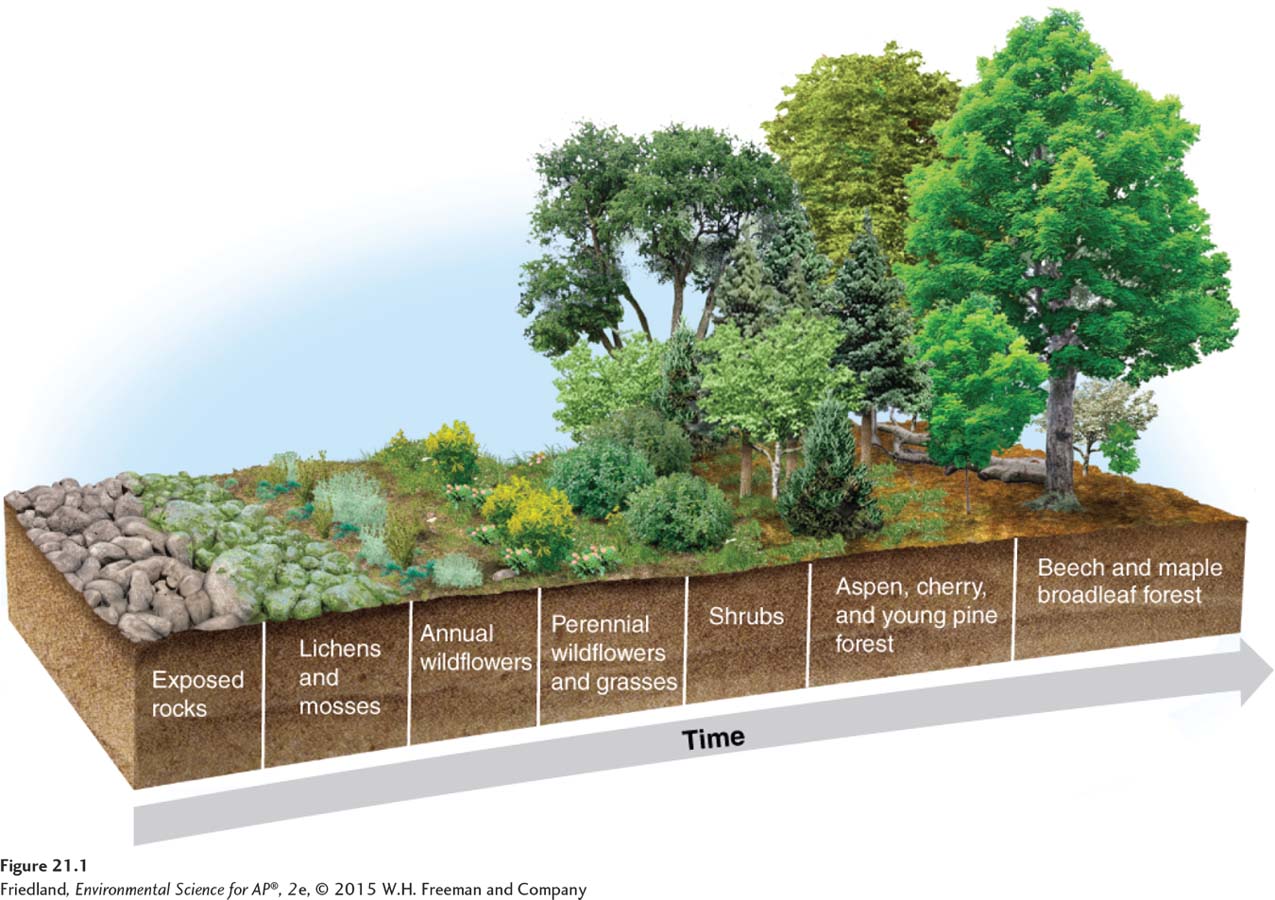
Over time, soil develops on the bare surface and it becomes a hospitable environment for plants with deep root systems. Mid-
The type of community that eventually develops is determined by the temperature and rainfall of the region. In the United States, succession produces forest communities in the East, grassland communities in the Midwest, and shrubland communities in the Southwest. In some areas, the number of species increases as succession proceeds. In others, late-
Secondary succession starts with soil
Secondary succession The succession of plant life that occurs in areas that have been disturbed but have not lost their soil.
Secondary succession occurs in areas that have been disturbed but have not lost their soil. Secondary succession follows an event, such as a forest fire or hurricane, that removes vegetation but leaves the soil mostly intact. Secondary succession also occurs on abandoned agricultural fields, such as the New England farms we discussed at the beginning of the chapter.
Pioneer species A species that can colonize new areas rapidly and grow well in full sunshine.
FIGURE 21.2 shows the process of secondary succession, again for a typical forest in New England. It usually begins with rapid colonization by plants that can easily disperse to the disturbed area. The first to arrive are typically such plants as grasses and wildflowers that have light, wind-
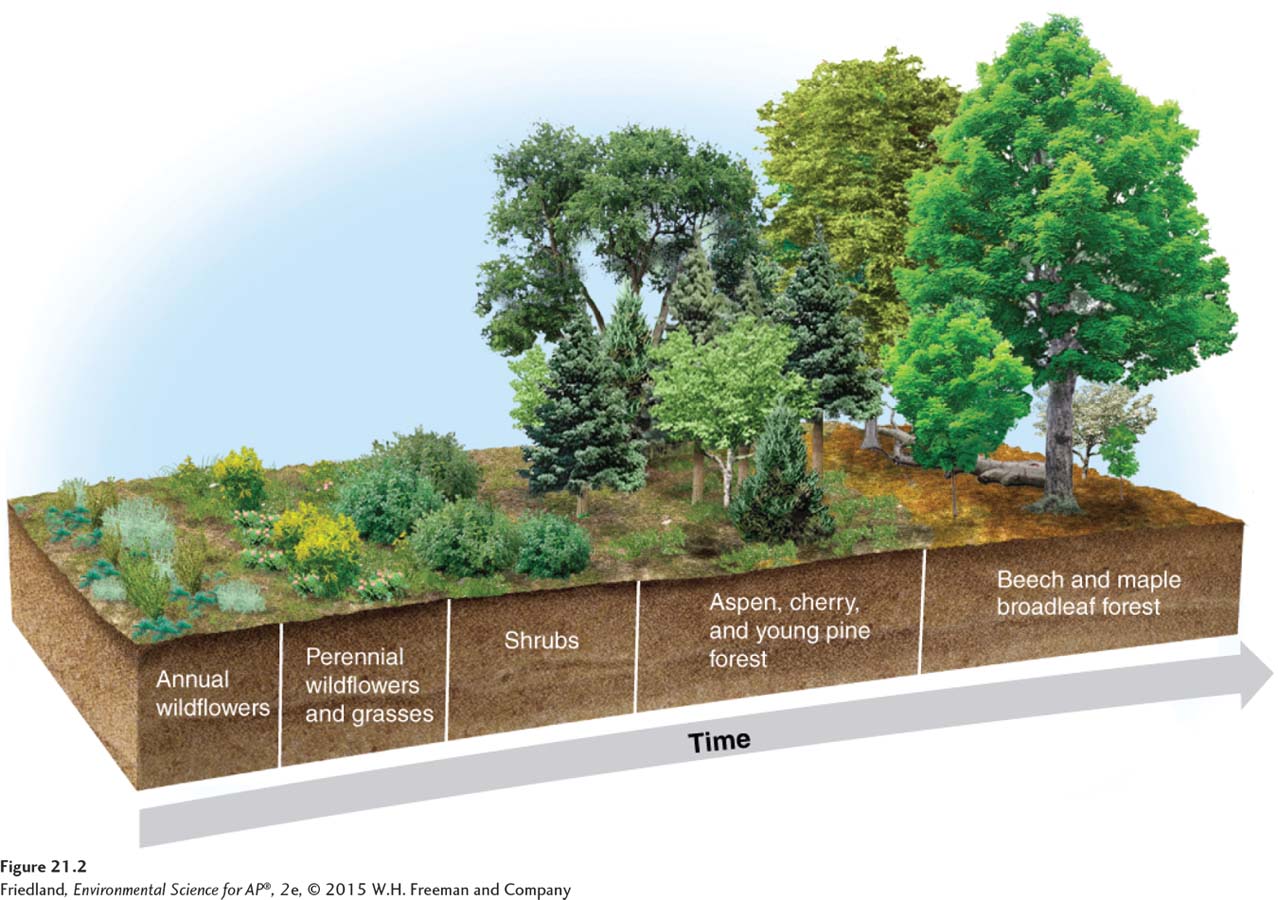
The process of secondary succession happens on many spatial scales, from small-
Historically, ecologists have described succession as ending with a climax stage. In forests, for example, ecologists considered the oldest forests to be climax forests. However, it is now recognized that, because natural disturbances such as fire, wind, and outbreaks of insect herbivores are a regular part of most communities, a late-
Succession occurs in a variety of aquatic ecosystems
Disturbances also create opportunities for succession in aquatic environments. For example, the rocky intertidal zone along the Pacific coast of North America is exposed to the air during low tide and lies under water during high tide. From time to time, major storms turn over rocks or clear their surfaces of living things. These bare rocks can then be colonized through the process of primary succession. Diatoms and short-
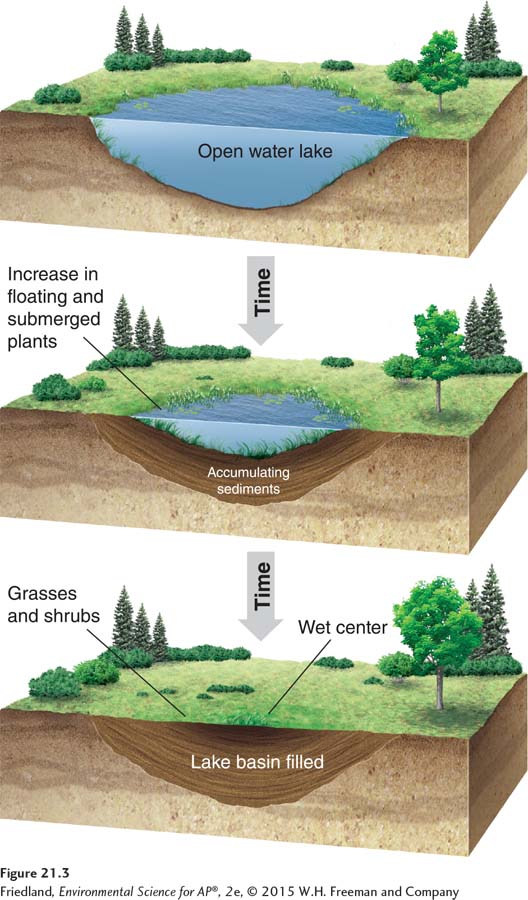
FIGURE 21.3 shows the pattern of succession in freshwater lakes. A glacier may carve out a lake basin, scouring it of sediments and vegetation. Over time, algae and aquatic plants colonize the lake. The growth of plants and algae and the erosion of surrounding rock and soil slowly fill the basin with sediment and organic matter, making it increasingly shallow. This process, which may take hundreds or even thousands of years, will eventually fill in the lake completely and make it a terrestrial habitat.
The species richness of a community is influenced by many factors
As we have seen, species are not distributed evenly on Earth. They are organized into biomes by global climate patterns and into communities whose composition changes regularly as species interact. In a given region within a biome, the number and types of species present are determined by three basic processes: colonization of the area by new species, speciation within the area, and losses from the area by extinction. The relative importance of these processes varies from region to region and is influenced by four factors: latitude, time, habitat size, and distance from other communities.
Latitude
As we move from the equator toward the North or South Pole, the number of species declines. For example, the southern latitudes of the United States support more than 12,000 species of plants, whereas a similar-
Time
Patterns of species richness are also regulated by time. The longer a habitat exists, the more colonization, speciation, and extinction can occur in that habitat. For example, Lake Baikal in Siberia—
Habitat size and distance from a source of species
Theory of island biogeography A theory that demonstrates the dual importance of habitat size and distance in determining species richness.
The final two factors that influence species richness are the size of the habitat and the distance of that habitat from a source of colonizing species. These factors are the basis for the theory of island biogeography, which demonstrates the dual importance of habitat size and distance from a mainland in determining species richness.
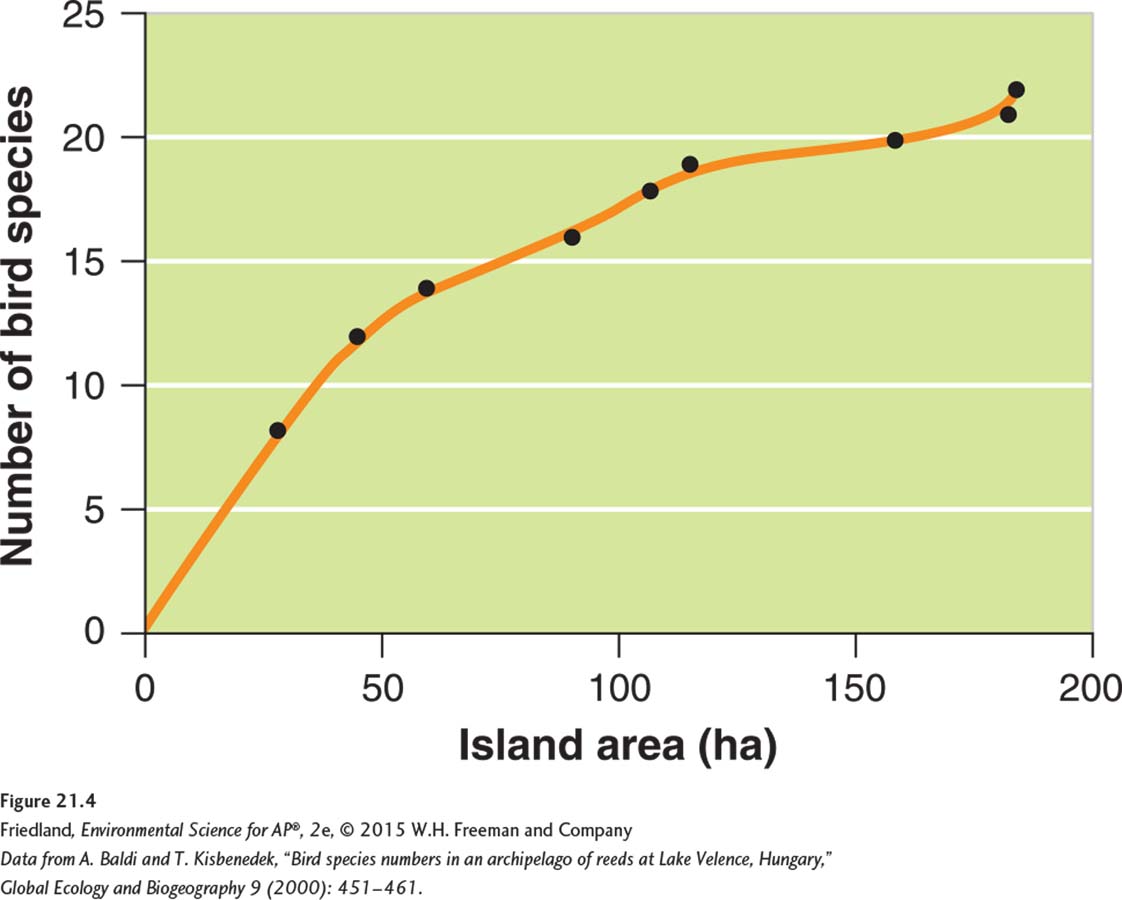
Larger habitats typically contain more species. In FIGURE 21.4, for example, we can see that larger islands of reed habitat in Hungary contain a greater number of bird species than smaller islands. There are three reasons for this pattern. First, dispersing species are more likely to find larger habitats than smaller habitats, particularly when those habitats are islands. Second, at any given latitude, larger habitats can support more species than smaller habitats. Larger habitats are capable of supporting larger populations of any given species, and larger populations are less prone to extinction. Third, larger habitats often contain a wider range of environmental conditions, which in turn provide more niches that support a larger number of species. A wider range of environmental conditions also provides greater opportunities for speciation over time.
The distance between a habitat and a source of colonizing species is the second factor that affects the species richness of communities. For example, oceanic islands that are more distant from continents generally have fewer species than islands that are closer to continents. Distance matters because, while many species can disperse short distances, only a few can disperse long distances. In other words, if two islands are the same size and contain the same resources, the nearer island should accumulate more species than the farther island because it has a higher rate of immigration by new species.
Conservation and island biogeography
The effects of colonization, speciation, and extinction on the species richness of communities have important implications for conservation. The theory of island biogeography was originally applied to oceanic islands, but it has since been applied to “habitat islands” within a continent, such as the “islands” of protected habitat represented by national parks. These habitat islands are often surrounded by less hospitable habitats that have been dramatically altered by human activities. If we wish to set aside natural habitat for a given species or a group of species, we need to consider both the size of the protected area and the distance between the protected area and other areas that could provide colonists.