4.3 Beyond Classical and Operant Theories of Learning: Play, Exploration, and Observation

Classical and operant conditioning have long been popular research topics in psychology, partly because they truly are ubiquitous learning processes and partly because they are relatively easy to study in the laboratory. By tightly controlling an animal’s environment and motivational state, a researcher can, more or less at will, induce predictable changes in the animal’s behavior. Indeed, the very word conditioning implies an active researcher (or trainer) and a passive animal. The researcher conditions the animal. The idea that learning is imposed by external conditions rather than controlled by the learner accords closely with the philosophy of early behaviorism.
127
In nature, however, animals (especially mammals) are active learners. They move about in ways that are well designed for learning. Through play, young animals learn how to control their own behavior in effective ways. Through exploration, animals of all ages keep track of significant changes in their environment. In addition, at least some animals acquire useful information through observing the behavior of others of their kind. Play, exploration, and observation are species-typical behavioral tendencies, or drives, that came about through natural selection precisely because they promote learning. Unlike operant conditioning as studied in laboratories, these activities occur most fully when the animal is free of any strong, immediate need state, such as hunger or thirst, and is free to move about at will.
Play: How the Young Learn How
Play is behavior that serves no obvious immediately useful purpose. It is behavior engaged in apparently for its own sake. The demeanor of the playing animal, just like that of a playing child, is often high-spirited and bouncing, which suggests that the animal is having fun. The young of all social mammals spend considerable portions of their time playing. Play is clearly species-typical: Nobody has to teach them to play; they just do it.
Why do young mammals play? It is not enough to say that they play because play is fun. That answer only begs us to ask, “Why are young mammals constructed in such a manner that they enjoy playing?” Natural selection does not create species-typical behaviors or pleasures that serve no purpose. So, what is the purpose of play?
Play clearly has some costs. It uses energy, which must be made up for in additional food, and it is sometimes directly dangerous. Young chimpanzees playfully chasing one another in trees sometimes fall and hurt themselves; goat kids frolicking on cliffs have been observed to slip and fall to their deaths; young fur seal pups are more likely to be snatched and eaten by predatory sea lions when they are at play than when they are at rest; and the play of cheetah cubs sometimes spoils their mothers’ attempts to stalk and capture game (Bjorklund & Pellegrini, 2002; Byers, 1998; Caro, 1995). The value of play must outweigh such costs, or natural selection would have weeded it out. What is that value?
Groos’s Theory: Play Is Practice of Species-Typical Skills
30
What is Groos’s theory about the evolutionary function of animals’ play, and what are five lines of evidence supporting that theory?
The first theorist to write about play from an evolutionary perspective was the German philosopher and naturalist Karl Groos. In a book entitled The Play of Animals, published in 1898, Groos argued that the primary purpose of play is to provide a means for young animals to practice their instincts—their species-typical behaviors. Groos was strongly influenced by the writings of Charles Darwin and had a sophisticated, modern understanding of instincts. He recognized that animals, especially mammals, must to varying degrees learn to use their instincts. Young mammals come into the world with biological drives and tendencies to behave in certain ways, but to be effective such behaviors must be practiced and refined. Play, according to Groos, provides that practice. This theory, even today, is the most widely accepted explanation of play among people who study it in animals.
Evidence for Groos’s Theory
Much of what we know about play in animals makes sense in the light of Groos’s theory and thereby provides evidence supporting that theory. Here are five categories of such evidence:
128
- Young animals play more than do adults of their species. Among the most obvious evidence for Groos’s practice theory is the simple fact that young animals, of all species that play, are much more playful than their elders. Young animals have more to learn than do adults of their species, so the fact that they are motivated to play more is no coincidence according to the practice theory.
- Species of animals that have the most to learn play the most. Another accurate prediction of Groos’s practice theory is that those animal species that have the most to learn play the most (Power, 2000). Young mammals play more than do the young of any other animal class, and mammals, more than any other class, depend on learning in order to survive. Among mammals, primates (monkeys and apes) are the most flexible and adaptable order, and the most dependent on learning; they are also the most playful of all animal orders. Among primates, human beings, chimpanzees, and bonobos are the most flexible and adaptable species, and they appear to be the most playful of all species. Also among mammals, those in the carnivore order (which includes the dog-like and catlike species) are more playful than herbivores, which is consistent with the observation that success in hunting requires more practice than does success in grazing.
- Young animals play most at those skills that they most need to learn. To a considerable degree it is possible to predict the activities that a young animal will play at by knowing what skills it must develop to survive and thrive. Young carnivores play at chasing, stalking, and pouncing—skills they will need in obtaining food (Schaller, 1972). Young herbivores do not play at grazing; that skill is easy to develop and can be practiced through serious grazing, so play is not needed. But young herbivores do play at fleeing and dodging, skills they need for evading predators (Spinka et al., 2001). The young of almost all mammals play at fighting. Among most species of mammals, young males playfight more than do young females (Pellegrini & Smith, 1998), and this corresponds with the fact that among most mammals fighting is a more essential adult skill for males than it is for females. At least among some species of primates, young females, but not young males, engage in playful care of infants (Maestripieri & Roney, 2006).
- Play involves much repetition. To practice something is to do it over and over again, and repetition is one of the hallmarks of play. Think of the difference between a cat seriously preying on a mouse and a cat playing at preying on a mouse. The preying cat pounces once, in its most efficient manner; it then kills the mouse and either eats it or takes it to its den. The playing cat, in contrast, lets the mouse go after catching it so it can stalk and pounce again—and again, and again, until the poor mouse dies of fright and exhaustion. Similarly, bear cubs playing chase games repeatedly chase one another, taking turns being the pursued and the pursuer, like children playing tag. And monkeys, playing at swinging from branch to branch in a tree, sometimes swing repeatedly between the same two branches (Symons, 1978). The repetition is not stereotyped; each repetition may be a little different from the previous one. It is as if the young animal is trying out, each time, a different way of preying, chasing, or swinging.
- Play is challenging. In play, young animals seem deliberately to put themselves into situations that challenge their abilities. In playfighting, whether among rats or monkeys, the stronger of the two will allow the weaker one to pin it and then will struggle to get out of the pinned position. In playful leaping and running, the young of many species seem to make deliberately awkward motions, from which they then have to recover skillfully to avoid a fall (Spinka et al., 2001). When young monkeys practice swinging from branch to branch, they commonly choose branches that are at such a distance that they often can’t make it, but low enough that a fall doesn’t hurt. Mountain goat kids that can already run well on flat ground tend to concentrate their playful running on steep slopes, where running is more difficult (Byers, 1977).
129
Applying the Theory to Humans
In a second book, entitled The Play of Man (1901), Groos extended his theory of play to human beings. Groos recognized basic differences between humans and other mammals. We humans have at least as many species-typical behaviors as other mammals have, but ours are even less rigid, more modifiable by experience, enabling us to adapt to a wider range of environmental niches than is true of other mammals. Moreover, we are the only truly cultural species. Culture, by definition, is the set of learned skills, knowledge, beliefs, and values that characterize a group of interconnected individuals and are passed along from generation to generation. To do well as human beings we must learn not just the skills that are common to the whole species, but also those that are unique to the specific culture in which we are developing.
So, according to Groos, in the evolution of our species the basic mammalian drive to play was elaborated upon, by natural selection, to include a heavy component of imitation. We are motivated to play not just at those activities (such as two-legged walking and running) that people everywhere do in pretty much the same way, but also to play at the very specific activities that we see are crucial to success in the particular culture in which we are growing up. Children in a hunting culture play at hunting, using and elaborating upon the particular methods of hunting that they see among their elders. Children in our present-day culture play at writing and computers. In the evolution of human beings, those individuals who were born with a genetic tendency to pay attention to and play at the human activities around them, especially those activities that are difficult yet crucial to success, were more likely to survive and reproduce than those born without such a tendency.
For Groos, and for most theorists since, play has been seen as a preparation for adulthood, and surely it is. But play also provides some immediate benefits for the players, be they human or nonhuman (Bjorklund & Green, 1992; Pellegrini, 2013). Some aspects of play serve to adapt children to the niche of childhood and not necessarily (or only) to prepare them for adulthood. Rough-and-tumble play provides opportunities for vigorous physical exercise that is important for skeletal and muscle development (Bruner, 1972; Dolhinow & Bishop, 1970). Children can also develop a sense of mastery during play when experimenting with new activities, including tool use, which can have benefits both for their immediate survival and for their future (Bjorklund & Gardiner, 2011). Moreover, the rough-and-tumble play characteristic of young males serves as a way for them to learn and practice social signaling (Martin & Caro, 1985), with exaggerated movements and a play face communicating playful intent. Males also use rough-and-tumble play to establish leadership in their peer group and assess others’ strength (Pellegrini & Smith, 1998), both important in dealing with one’s peers in the here and now, not just in some unspecified future.
Exploration: How Animals Learn What and Where
31
How does exploration differ from play in its evolutionary function?
Play is not the only drive that came about in evolution to promote active learning. The other great one is curiosity, or the drive to explore. Groos (1898) considered exploration to be a category of play, but most students of play and exploration now consider the two to be distinct (Pellegrini, 2013). Learning can be divided at least roughly into two broad categories—learning to do (skill learning) and learning about (information learning). Play evolved to serve the former, and exploration evolved to serve the latter.

Exploration is a more primitive and widespread category of behavior than is play. Animals whose activities are rather rigidly controlled by their genetic makeup so that they don’t have much to learn in the to do category must nevertheless learn about their environment. They must learn where food, shelter, mates, and other necessities for life and reproduction are located. Fish don’t play, at least not in any way that is reliably identified as such by researchers, but they do regularly approach and explore novel objects (Ward et al., 2003). Even insects explore to find food and other necessities of life, although they do so in rather inflexible ways (Gordon, 1995). Mammals of all species, regardless of age, explore novel environments and objects with which they are confronted.
130
The Nature of Mammalian Exploration

Exploration, unlike play, is often mixed with a degree of fear. Exploration is elicited by novel stimuli, and novel stimuli often induce fear until they are fully explored. In fact, one purpose of exploration, in animals and people, is to determine whether or not an unfamiliar object or place is safe. Explorers are often caught in a balance between curiosity, which drives them toward the unfamiliar terrain or novel object, and fear, which drives them away. A rat placed in a novel test arena with various objects in it will first cower in a corner. Then it will take a few steps out from the corner, along a wall, and dash back. Then it will venture a bit farther along the wall before dashing back again. Gradually, the rat will get bolder. It will eventually approach and explore—by smell, sight, and touch—the entire arena and all the objects within it. Once the rat is thoroughly familiar with the arena, it will reduce its movement but will continue periodically to tour the arena as if looking to see if anything has changed—a behavior referred to as patrolling. During its patrolling, the animal periodically rears up on its hind legs to get a better view. If a new object has been placed in the arena, the rat will attend to that rather than to old objects and will explore it at first gingerly and then more boldly (Inglis et al., 2001; Renner & Seltzer, 1991). Similar behaviors have been described in many other species of mammals.
Some of the earliest research on exploration came from studies of rats in mazes. Researchers learned that rats’ movements through mazes are governed not just by their drive for the food in the goal box, but also by their drive to explore all of the maze’s alleys (Dember & Fowler, 1958; Winfield & Dennis, 1934). Rats that have already learned the most direct route to the goal will often persist in exploring roundabout routes and dead-end alleys. Rats that are prevented from entering a particular alley, by a block placed at its entrance, will typically explore that alley first as soon as the block is removed. Not surprisingly, the hungrier a rat is (that is, the longer it has been deprived of food), the more directly it will run to the food-containing goal box; the less hungry it is, the more time it will spend exploring the other alleys in a maze. But even very hungry rats will often spend some time exploring (Inglis et al., 2001).
Evidence That Animals Acquire Useful Information Through Exploration
32
How do rats explore a novel environment? How did Tolman and subsequent researchers show that rats learn useful information in their exploration?
In a now classic experiment, Edward Tolman and C. H. Honzik (1930) showed that rats can learn about the pathways in a maze even if no food or other such reward is provided for doing so. These researchers tested three groups of rats in a complex maze under different reward conditions. Group 1 received one trial per day in the maze with no food or other reward in the goal box. As expected, this group showed little improvement from day to day in the time they took to reach the goal box (the goal box contained no “goal” for them). Group 2 received one trial per day with food in the goal box. As expected, this group improved considerably from day to day in their rate of movement to the goal box. The most interesting group was group 3. Rats in this group received one trial per day with no reward for 10 days, like group 1; but, beginning on the 11th day, they received one trial per day with a food reward, like group 2. These rats improved dramatically between days 11 and 12. On day 11, they were no better than the other unrewarded group (group 1), but on day 12, after just one experience with the reward, they were as fast at reaching the goal box as the rats that had been rewarded all along (see Figure 4.15).
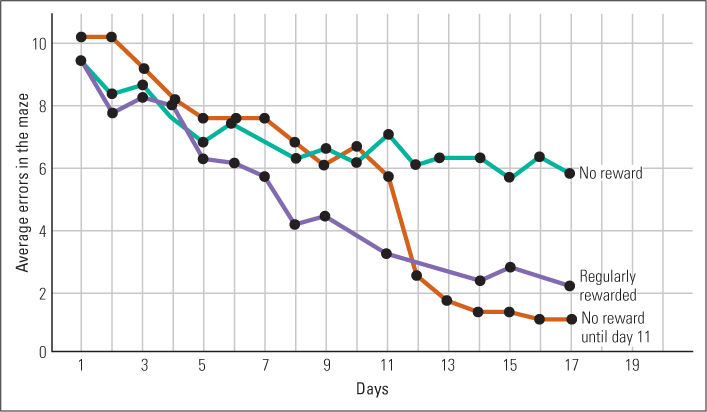
131
On the basis of this and other experiments, Tolman (1948) argued that rewards affect what animals do more than what they learn. Animals learn the locations of distinctive places in their environment through exploration, whether or not they have ever found rewards there, but they do not run directly to those places unless they have found rewards there. Tolman used the term latent learning to refer to learning that is not immediately demonstrated in the animal’s behavior. In the experiment just described, the rats in group 3 learned the spatial layout of the maze in the first 10 trials, but that learning remained latent, not manifested in their behavior, until the addition of a reward gave the rats a reason to run straight to the goal box.
Latent learning has been demonstrated in many experiments since Tolman’s time. Through exploration, without rewards, rats learn the layouts of mazes and then are able to take the shortest route to a reward when a reward is introduced (Roberts et al., 2007). Animals also learn about potential hiding places through exploration (Leonard & McNaughton, 1990). In one set of experiments, rats that had an opportunity to explore a novel arena that contained one or more hiding places ran much more quickly to a hiding place, when they were deliberately frightened in a later test, than did rats that had not previously explored the arena (Renner, 1988). There is also evidence, from experiments with mice, that animals that explore the most are the same animals that learn the most in a wide variety of tests of learning (Matzel et al., 2006).
In nature it is likely that most learning about the environment is latent. Animals learn about many aspects of their surroundings through exploration, but only an unpredictable portion of that knowledge becomes useful at a later time and affects the animal’s behavior.
Social Learning: Learning by Watching and Interacting with Others
The objects of most intense exploration, for most people, are other people. By watching other people we learn an enormous amount about people, about how to do things, and about the human significance of the rest of our environment. Social learning is broadly defined as occurring in a situation “in which one individual comes to behave similarly to another” (Boesch & Tomasello, 1998). This is usually done simply by watching others and is referred to by psychologists as observational learning.
An early (and continuing) advocate of the importance of observational learning in humans is Albert Bandura (1962, 1986). In his social-learning (later renamed social-cognitive) theory, Bandura argued that children learn important social behaviors from observing others through a process of vicarious reinforcement, the ability to learn from the consequences of others’ actions. It is not necessary for a person’s behavior to be explicitly reinforced to learn; rather, it is enough to observe another person reinforced for some action. As Bandura modified his theory over nearly 50 years, his outlook became increasingly more cognitive, deviating from its original roots in behaviorism. For example, in addition to vicarius reinforcement, Bandura proposed four other key cognitive capabilities—symbolization, forethought, self-regulation, and self-selection—which are described in Table 4.1.
Key cognitive capabilities in Bandura’s social-cognitive theory

132
Through observation we learn about the unique characteristics of others, so we can judge how to get along with them and know who is likely to help us or hurt us in a given situation. When we are in a new social environment, we look frequently to others to learn what sorts of behaviors are normal or expected in that setting—so when in Rome, we can do as the Romans do. We also look frequently to others to see what they are looking at and how they are responding to what they are looking at. In that way we learn some of what they already know about the surrounding environment. To learn a new skill—whether it’s a new dance step, driving a car, or surgery—we usually begin by observing the actions of a person who has already mastered the skill. Thank goodness for that; imagine what the world would be like if people learned driving and surgery only by trial and error!
Learning How by Watching Skilled Performers
33
How does observation of skilled performers help animals learn new operant tasks? How does imitation differ from stimulus enhancement, goal enhancement, and emulation? What evidence suggests that primates (especially chimpanzees), but not other mammals, are capable of imitation?
Many experiments have shown that animals can learn or partially learn how to perform a new task by watching others do it. Kittens learned more quickly to press a lever for food if they had seen their mother do so than if they had not (Chesler, 1969). Dogs were better at moving around a barrier to get food if they had seen another dog or human being do it first than if they hadn’t (Pongrácz et al., 2008). Mice who had observed a well-trained mouse make a sequence of two movements to get a food pellet learned that sequence of movements more readily than those who hadn’t (Carlier & Jamon, 2006).
It is tempting to interpret such results as evidence that mammals in general are able to learn by imitating the actions of others of their kind. But, if you think about it, you realize that imitation is cognitively complex. To imitate, an animal must observe, remember, and reproduce the specific pattern of movements that were produced by the model. To reproduce the movements, the learner must map the observed actions onto its own movement control system. Many researchers argue, with evidence, that real imitation does not occur in mammals other than primates (and perhaps only in humans; Bjorklund et al., 2010; Whiten et al., 2004). Further, they point to evidence that observational learning by nonprimates involves simpler means, including stimulus enhancement and goal enhancement (Byrne & Russon, 1988; Zentall, 2003, 2006).
133
Stimulus enhancement refers to an increase in the salience or attractiveness of the object that the observed individual is acting upon. Goal enhancement refers to an increased drive to obtain rewards similar to what the observed individual is receiving. Thus, a kitten that sees its mother pressing a lever for food pellets may become attracted to the lever (stimulus enhancement) and motivated to eat food pellets (goal enhancement). In this case, stimulus enhancement increases the likelihood that the kitten will press the lever, goal enhancement increases the reward value of the pellets, and the two combined help the kitten learn very quickly to press the lever for pellets. By this account, the kitten is not imitating the movements of its mother but is going to the lever because that was made salient, pushing it more or less accidentally, and then eating the pellets because they too were made salient.
Chimpanzees clearly learn by observation, but they seem not to engage in true imitation, at least not much. Rather, they are more apt to learn by emulation, which involves observing another individual achieve some goal (picking up and dropping a log to reveal tasty ants, for instance), then reaching that same goal by their own means (bouncing up and down on the log). For example, in one study, 3- and 4-year-old children and chimpanzees were shown a series of three actions, two of which were necessary and one of which was not, to open an opaque puzzle box. Both the children and the chimpanzees copied all three behaviors and got the reward. However, when they were then shown a transparent puzzle box, the children still copied all three actions, even when they could readily see that one of these actions was irrelevant. The chimpanzees, however, skipped the irrelevant action and copied only the necessary ones to achieve their goal (Horner & Whiten, 2005). In this and other studies (Nielsen, 2012), the chimpanzees seem to be more focused on the goal and less on the means used to achieve it, a characteristic of emulation.
There is at least one exception to this pattern, and that is for chimpanzees that have been raised as if there were children, called enculturated chimpanzees. In several studies, such animals display true imitation, even imitating the behaviors of a model up to 24 hours after originally observing it (Bjorklund et al., 2002; Buttlemann et al., 2007; Tomasello et al., 1993). Apparently, something associated with a humanlike rearing environment altered young chimpanzees’ social-learning abilities, causing them to be more like 4-year-old human children in some ways than mother-raised chimpanzees.
Neuroscientists have discovered that the brains of human beings and of at least some nonhuman primates contain organized systems of neurons (nerve cells) that seem to be well designed to make imitation easy and natural (Iacoboni & Dapretto, 2006; Jaffe, 2007). The neurons are referred to as mirror neurons because they are believed to help us behave in ways that mirror (mimic) what we see. Within this system, the same neurons that become active when the individual makes a particular motion also become active when the individual sees another individual make that motion. For example, the same neurons that are exercised when making an overhand serve in tennis are also exercised when watching someone else make an overhand serve. In terms of its effect on these neurons, observing is like doing; thus, observing an action may alter the neurons and allow for new learning through the same means that occur when performing the action.
Cultural Transmission in Chimpanzees
34
What is the evidence that chimpanzees transmit cultural traditions from generation to generation?
We humans are the supremely cultural animal. Culture, as we said before, refers to the beliefs and traditions that are passed along from generation to generation. Because of culture we do not have to reinvent the wheel, or figure out how to grow crops or how to build houses; we merely improve on what our ancestors invented. Observational learning appears to be a prerequisite for human culture. The skills and rituals acquired by each generation are passed on to the next not so much because the older generation deliberately trains the younger (though that is part of it) but more because members of the younger generation intently observe their elders and learn to behave as they do.
134

The species that comes second to us in exhibition of culture—though a far distant second—is, not surprisingly, the chimpanzee. Wild chimpanzees living in different groups, geographically isolated from one another, have different cultural traditions, which pass from generation to generation. Researchers studying wild chimpanzees at seven different field stations have identified at least 39 different behaviors, ranging from tool design to mating displays, that are distinct to specific groups and that seem to arise from cultural tradition rather than from constraints imposed by the environment (Whiten, 2007; Whiten et al., 1999).
For example, chimpanzees in some colonies crack hard-shelled nuts by placing a nut on a carefully selected rock and hitting it with another carefully selected rock (see Figure 4.16; Mercader et al., 2002). The young in these colonies learn this skill by observing their elders and practicing elements of it over a period of several months (Inoue-Nakamura & Matsuzawa, 1997). In other colonies, however, chimpanzees have never been observed to crack nuts, even though plenty of nuts and stones are available. Apparently, in those colonies the discovery that nuts can be cracked was never made, so it could not be passed along. Research conducted with chimpanzees living in a wildlife sanctuary has shown that the introduction of one nut-cracking chimp can lead quickly to the spreading of that skill throughout the colony (Marshall-Pescini & Whiten, 2008).
The most sophisticated form of social learning is teaching, which involves the teacher modifying his or her behavior in order that the “student” acquire new knowledge. Teaching requires that the learner appreciate the perspective of the teacher and that the teacher be sensitive to the knowledge, motivations, and emotions of the learner. Parents begin “teaching” their children in infancy, although true learning from teaching likely doesn’t occur until sometime later. Although we frequently talk of nonhuman animals teaching their offspring, teaching in the animal world is rare. It has been shown in meerkats (Thorton & McAuliffe, 2006), dolphins (Bender et al., 2009), and chimpanzees (Boesch, 1991; Greenfield et al., 2000), but the situations in which it is observed are far more limited than in humans. For example, when mother Atlantic spotted dolphins are foraging with their calves, they orient their bodies to the prey they are chasing and delay eating the prey (Bender et al., 2009). Mother chimpanzees make exaggerated movements while cracking nuts when in the presence of their infants (Boesch, 1991). However, at least in chimpanzees, teaching is rarely observed, and it appears that most social learning in our close genetic relatives is not achieved by teaching but by other less complex forms observational learning, such as emulation (Bering & Povinelli, 2003).
Gaze Following as an Aid in Learning from Others
35
How might gaze following help us learn from other people? What characteristic of human eyes makes gaze following easier for us than for other primates?
When we are attending to another person our eyes move, automatically, reflexively, in the same direction that his or her eyes move, so we look at the same object at which he or she is looking. This reflexive action, called gaze following, helps us understand what the other person is thinking about and, in conversations, helps us to know what he or she is talking about (Richardson et al., 2007). When a person says, “Oh, that is beautiful,” our gaze following helps us figure out what that refers to.
135

Gaze following has been most extensively studied in young children. Beginning in the latter half of their first year of life, human infants reflexively tend to look at whatever their caregiver is looking at (Brooks & Meltzoff, 2002; Woodward, 2003). Such gaze following ensures that the infants pay attention to the same objects and events in their environment that their elders attend to, which may be the most important things to learn about for survival in their culture. It also, no doubt, helps infants learn language. If you are an infant and hear your mother say some new word—maybe goat—for the first time, you have a chance of learning what that word refers to if you are looking at the same object that she is looking at. Consistent with this idea, researchers have found that babies who show the most reliable gaze following learn language faster than those who exhibit less gaze following (Brooks & Meltzoff, 2008).
No other animal follows the gaze of another to the extent that we humans do. In fact, the unique coloring of our eyes may be a special human adaptation that came about through natural selection to enable us to follow each others’ gazes and thereby understand each other better (Tomasello, 2007). The relatively dark circular iris of the human eye is sharply set off by the bright white of the rest of the visible portion of the eyeball, the sclera, which makes it easy for others to see where we are looking (see Figure 4.17). Other primates, including chimpanzees and bonobos, have dark sclera, which do not contrast with the iris, so it is not possible to see which way their eyes have shifted (look back at the nut-cracking chimpanzee in Figure 4.15). Research has shown that chimpanzees and bonobos do engage in some gaze following and are able to learn through that means, but their gaze following is less automatic than humans’ and depends entirely on observations of the model’s head, not on shifts of the eyes within their sockets (Okamoto-Barth et al., 2007; Tomonaga, 2007).
Of course, one disadvantage of our white scleras is that it is hard for us to deceive others about where we are looking. For that, we need sunglasses. People wearing sunglasses seem a little intimidating to us, partly because we don’t know where they are looking, which reduces our ability to guess what they are thinking. Presumably, in the course of our evolution, the advantages of cuing others into our thoughts outweighed the advantages of deception.
As we’ve seen, many species engage in social learning, but none does so to the extent and level of proficiency as Homo sapiens. Many have suggested that human minds were shaped by natural selection to be especially attentive to social relations—to infer the goals, intentions, and thoughts of others—that permitted our ancestors to more effectively cooperate and compete with, and especially learn from one another (Shafto et al., 2012; Tomasello, 2009). You can think of humans’ propensity for social learning as a broad type of specialized learning mechanism, and we will examine the centrality of social cognition for our species in later chapters of this book. But now we turn to some narrower forms of specialized learning, both in humans and other animals.
SECTION REVIEW
In the natural environment, learning is promoted by play, exploration, and observation.
Play
- In Line with a theory originated by Karl Groos, young mammals appear to play in ways that help them to develop crucial survival skills. Play is most frequent in those species that have the most to learn.
- Human children play not just at skills that are crucial to people everywhere, but also at those unique skills that are crucial to the culture in which they develop.
Exploration
- Exploration is more primitive than play, occurring in many more species and at all ages. It promotes learning about the environment.
- Curiosity motivates exploration of novel objects and places, but is balanced by fear.
- Latent learning experiments show that exploration alone, without external reward, produces useful knowledge.
Social Learning
- Albert Bandura’s social-learning (later social-cognitive) theory emphasized the role of vicarious reinforcement in social learning.
- Stimulus enhancement, goal enhancement, and emulation are simpler forms of observational learning than imitation. Mirror neurons may facilitate social learning in primates.
- The most sophisticated form of social learning is teaching, which humans do readily but is rare (though not nonexistent) on other animals.
- Reflexive gaze following helps humans learn from one another
136