6.5 The Sleep Drive
Sleepiness is clearly a drive. A sleepy person is motivated to go to sleep and will expend effort to reach a safe and comfortable place to do so. Achieving this goal and drifting off to sleep provides a sense of pleasure analogous to that which comes from eating when hungry or copulating when sexually motivated.
221
Sleepiness operates in some ways like a regulatory drive. As with hunger, thirst, or other regulatory drives, the longer one goes without satisfying the sleep drive, the stronger the drive becomes. But, unlike other regulatory drives, it is not clear what the sleep drive regulates, except sleep itself. In addition, sleepiness is controlled not just by amount of sleep deprivation but also by a biological clock that keeps time within the 24-hour day–night cycle. Regardless of how much sleep one has had recently, sleepiness tends to increase at night and decrease during daytime hours. The discussion in this section focuses on three questions: (1) What is sleep? (2) What are the functions of sleep? (3) What brain mechanisms control sleepiness and arousal?
Description of Sleep as a Physiological and Behavioral State
Sleep is a condition of relative unresponsiveness to the environment. Because people show little overt behavior and cannot answer questions when they are asleep, scientists who study this state must focus on physiological and subtle behavioral changes.
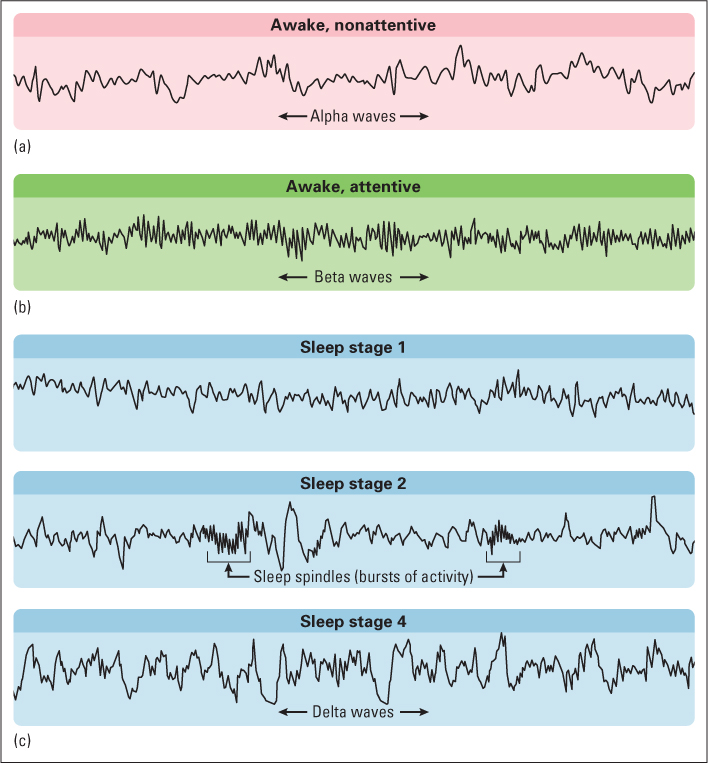
The most valuable index of sleep is based on the electroencephalogram (abbreviated EEG). As discussed in Chapter 5, the EEG is an amplified recording of the electrical activity of the brain that is picked up by electrodes pasted to the person’s skull. These electrical signals can be stored and analyzed by a computer, producing records like those shown in Figure 6.10. The EEG recording is a gross index of the electrical activity of the brain, representing a sort of average of the activity of billions of neurons, with the greatest weight given to those lying closest to the recording site.
222
EEG Waves Accompanying Wakefulness and Stages of Sleep
27
How does a person’s EEG change as the person goes from alert to relaxed to various stages of sleep?
When a person is relaxed but awake, with eyes closed and not thinking of anything in particular, the EEG typically consists of large, regular waves called alpha waves, which occur at a frequency of about 8 to 13 cycles per second (see Figure 6.10b). These relatively slow waves stem from a synchronized pulsing of neurons in the thalamus and cerebral cortex that occurs in the absence of focused mental activity or emotional excitement. When a person concentrates on an external stimulus, or tries to solve a problem, or becomes excited, the EEG pattern changes to low-amplitude, fast, irregular waves called beta waves (see Figure 6.10a). The low amplitude of these waves indicates that neurons are firing in an unsynchronized manner, such that their contributions to the EEG tend to cancel one another out. Whereas alpha waves are analogous to the large, regular waves that occur on a pond undisturbed by anything but a steady wind, beta waves are more akin to the effect of a million pebbles tossed suddenly onto the surface of the pond. The crests of the ripples created by some pebbles would cancel out the troughs created by others, resulting in a chaotic, high-frequency, low-amplitude pattern of ripples.

When a person falls asleep, the EEG goes through a fairly regular sequence of changes, which are used by researchers to divide sleep into four stages, using the criteria described in the caption to Figure 6.10. Stage 1 is a brief transition stage, when the person is first falling asleep, and stages 2 through 4 are successively deeper stages of true sleep. As sleep deepens, an increased percentage of the EEG is devoted to slow, irregular, high-amplitude waves called delta waves. These waves, like others, are controlled by neurons in the thalamus that respond in an oscillating manner and synchronize the activity of billions of neurons in the cerebral cortex (Steriade et al., 1993). Corresponding to this EEG change, muscle tension, heart rate, and breathing rate decline, and the person becomes increasingly hard to awaken.
Cyclic Repetition of Sleep Stages Through the Night
28
How do REM and non-REM sleep differ, and how do they cycle through the night?
Having reached stage 4, a person does not remain there for the rest of the night. Instead, after about 80 to 100 minutes of total sleep time, sleep rapidly lightens, returning through stages 3 and 2, and then a new, quite fascinating stage of sleep appears for a period of about 10 minutes or more. During this new stage the EEG is unsynchronized, looking much like the beta waves of alert wakefulness. On the basis of the EEG alone, one might think that the person had awakened, but direct observation shows that the person is sound asleep, and the record of muscle tension shows that the muscles are more relaxed than at any other sleep stage. Yet, consistent with the unsynchronized EEG, other indices of high arousal are apparent: Breathing and heart rate become more rapid and less regular; penile erection occurs in males (even in infants and young boys); twitching movements occur in the small muscles of the fingers and face; and, most indicative of all, the eyes move rapidly back and forth and up and down under the closed eyelids. These eye movements, which can be recorded electrically along with the EEG, give this stage of sleep its name, rapid-eye-movement sleep, abbreviated REM sleep. As you may have guessed, it is during REM sleep that most dreams occur. REM sleep is also sometimes called emergent stage 1, because, even though it is different from the original stage 1, it marks the onset of a new sleep cycle. Stages 2, 3, and 4 are referred to collectively as non-REM sleep.
223
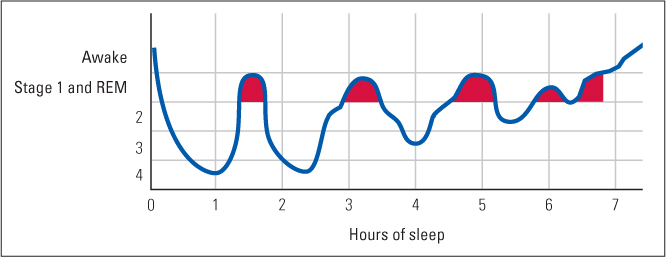
In a typical night’s sleep, a person goes through four or five sleep cycles, each involving gradual descent into deeper stages of non-REM sleep, followed by a rapid lightening of non-REM sleep, followed by REM sleep (Hobson, 1995). Each complete cycle takes about 90 minutes. As depicted in Figure 6.11, the deepest non-REM sleep occurs in the first cycle or two. With each successive cycle, less time is spent in the deeper stages of non-REM sleep (stages 3 and 4), and more time is spent in light non-REM sleep (stage 2) and REM sleep.
Dreams and Other Mental Activity During Sleep
29
What are some general characteristics of dreams that people describe when aroused from REM sleep, and how do these differ from the “sleep thought” that people more often describe when aroused from non-REM sleep?
When people are awakened during REM sleep, they usually (in about 90 percent of cases) report a mental experience that researchers call a true dream (Foulkes, 1985). Such a dream is experienced as if it were a real event rather than something merely imagined or thought about. The dreamer has the feeling of actually seeing or in other ways sensing various objects and people and of actually moving and behaving in the dream environment. Moreover, a true dream usually involves a progression of such experiences, woven into a somewhat coherent though often bizarre story. The more time the sleeper spends in REM sleep before awakening, the longer and more elaborate is the reported dream. Studies show that essentially everyone dreams several times a night. Those who believe that they rarely dream, or who can recall only fragments of dreams upon normal awakening in the morning, describe vivid, detailed dreams if awakened during REM periods.
Analyses of the contents of hundreds of reported dreams reveal a number of generalities about them (Domhoff, 2003; Valli et al., 2008). Most dreams are about people, objects, and activities that are well known and meaningful to the dreamer, but very few dreams are repetitions of events that actually happened in the dreamer’s daytime experience. Most dreams involve emotions, especially negative emotions. Dreams involving fear, worry, or embarrassment are more common than joyous dreams. Among college students, dreams of being lost, of being late for an examination, and of being inappropriately dressed (or undressed) in public are common.
People who are awakened during non-REM sleep report some sort of mental activity just before awakening roughly half the time (Foulkes, 1985; Hobson, 1995). Such reports are sometimes of true dreams, but more often they are of sleep thought, which lacks the vivid sensory and motor hallucinations of true dreams and is more akin to daytime thinking. Often the subject of sleep thought is some problem that had been of concern during the day. For example, a student who had been cramming for a math exam might report working on a calculus problem while sleeping. A major difference between sleep thought and daytime thought is that the former is usually ineffective. Although the sleeper may feel that he or she is solving a calculus problem, questions upon awakening indicate that no real progress was made (Hobson, 1987).
224

During sleep a person is less responsive to events in the environment than when awake, but not completely unresponsive. The eyes are closed, but all of the other sensory channels remain open. In one study using fMRI, people’s brains were observed to respond to various sounds when they were asleep in much the same way as they did when they were awake (Portas et al., 2000). Both when asleep and when awake, the sound of the person’s own name had effects on emotional centers in the limbic system that did not occur in response to sounds that were less meaningful to the person. The fact that parents can sleep through a thunderstorm but become aroused by the whimpering of their child in the next room is further evidence that the sleeping person’s brain sorts out sounds by meaning to some degree.
One characteristic of all mental activity during sleep is that it is quickly forgotten. Dreams, sleep thoughts, and sensory experiences during sleep are lost unless the person wakes up during them and thinks about them while awake. This is fortunate. Such forgetting prevents us from carrying around thousands of bizarre and confusing memories or of mixing up real events with dreamed ones.
Theories About the Functions of Sleep
Why must we sleep? Countless children have asked that question to protest being put to bed, and many scientists have asked it, too. Sleep must have come about in the course of evolution to serve some function or functions related to survival and reproduction; otherwise it would not be such a universal and compelling drive. Several theories have been proposed to explain the evolution of the sleep drive. The theories are not incompatible with one another, and they all appear to have some degree of validity.
The Preservation and Protection Theory
The preservation and protection theory of sleep derives primarily from comparison of sleep patterns across different species of animals. It posits that sleep came about in evolution to preserve energy and protect individuals during that portion of each 24-hour day when there is relatively little value and considerable danger in moving about. An animal needs only a certain number of hours per day to do the things that are necessary or useful for survival, and the rest of the time, according to this theory, it is better off asleep—quiet, hidden, and protected from predators and other possible dangers (Meddis, 1977).
225
30
What evidence supports the preservation and protection theory of sleep?
Support for this theory comes from evidence that variations in sleep time among different species do not correspond with differences in physical exertion while awake but do correspond with feeding habits and ways of achieving safety (Allison & Cicchetti, 1976; Lima et al., 2005). At one extreme, large grazing animals such as bison and horses average only 2 or 3 hours of sleep per 24-hour day. Because of their large size and because they eat grass and other vegetation, which are extremely low in calories, they must spend most of their time eating, and, therefore, they have little time to sleep. Moreover, because of their size and the fact that they cannot burrow or climb trees, such animals are not adept at finding safe places in which to sleep. Thus, they are safer awake.
Even among animals that are roughly the same size as each other, grazing animals sleep less than do meat-eaters. Sheep and goats, for example, sleep only 4 or 5 hours per 24 hours, while lions and tigers sleep 14 to 16 hours (Campbell & Tobler, 1984). Sheep and goats must spend more time eating than lions and tigers, and, because they are more often preyed upon, the former are at much greater risk when asleep than are the latter. At the extreme in sleep time are opossums and bats, which average about 20 hours of sleep each 24-hour day. These two species need little time to obtain food (such as high-calorie insects or grubs), and they are adapted to hide in out-of-the-way places. According to the preservation and protection theory, they sleep so much because they have no need to be awake for long and are protected from predators while asleep.

In addition to explaining differences in total amount of sleep, the preservation and protection theory also explains differences in the time of day at which different species sleep. Animals that rely heavily on vision generally forage during the day and sleep at night. Conversely, animals such as mice and rats that rely more on other senses, and are preyed upon by animals that use vision, generally sleep during the day and forage at night. The theory also offers an explanation for the fact that infants in most species of mammals sleep much more than adults. Infants who are being cared for by adults do not need to spend time foraging, and sleep protects them from wandering away into danger. Their sleep also gives their caregivers an opportunity to rest or attend to other needs.
It is interesting to speculate, in this vein, about the evolutionary conditions behind the 8-hour nighttime sleep pattern that characterizes adult humans throughout the world. Humans are highly visual creatures who need light to find food and do other things necessary for survival. At night it may have been best for us, during most of our evolution, to be tucked away asleep in a cave or other hiding place, so as not to be tempted to walk about and risk falling over a cliff or being attacked by a nocturnal predator. Only during the past few centuries—an insignificant speck of evolutionary time—have lights and other contrivances of civilization made the night relatively safe for us. According to this line of thinking, our pattern of sleep might be in part a vestigial trait, a carryover from a period when the night was a time of great danger. To the degree that nighttime is still more dangerous than daytime, our pattern of sleep may continue to serve an adaptive function.
The Body-Restoration Theory
The body-restoration theory of sleep function is the theory that most people intuitively believe. It is the theory that your parents probably repeated to you as their reason for sending you to bed at a certain hour. According to this view, the body wears out during the day, and sleep is necessary to put it back in shape.
226
31
What evidence supports the body restoration theory of sleep, and what are some limitations of the theory?
Scientific support for this theory includes the observation that sleep is a time of rest and recuperation. The muscles are relaxed, metabolic rate is down, and growth hormone, which promotes body repair, is secreted at a much higher rate than during wakefulness (Douglas, 2002; Siegel, 2003). Also consistent with the restoration theory is the observation that prolonged, complete sleep deprivation in rats results in breakdown of various bodily tissues, leading, within about 3 weeks, to death (Everson, 1993; Everson et al., 1989).
The theory also offers an explanation for the general tendency of small mammals to sleep longer than large ones. Small mammals need to maintain a higher overall level of metabolism than do large mammals because they lose body heat more rapidly, and higher metabolism leads to greater wear and tear on bodily tissues (Siegel, 2005). However, contrary to this theory, research with birds has failed to show any correlation across species between sleep time and metabolic rate; instead, it has shown a strong correlation between sleep time and risk of predation, which tends to support the preservation and protection theory discussed previously. Thus, birds that are most protected from predators while asleep, sleep longest regardless of their body size or metabolic rate (Roth et al., 2006). The theory also does not explain the large differences in sleep time between grazing animals and meat-eating animals that have similar body sizes and metabolic rates. Nor does it explain the failure of researchers to find consistent positive correlations, either across species or within species, between the amount of time an animal sleeps and the average amount of energy it expends through vigorous activity during the day.
The fact that all vertebrate animals sleep at least an hour or two out of every 24 hours, regardless of the degree to which they are at risk while sleeping, suggests that some amount of sleep is needed for body repair. But the body-restoration theory does not provide a satisfactory explanation of the large differences among species in sleep time, and it offers no explanation for the fact that some animals sleep during the day while others sleep at night. (Chapter 9 discusses an additional apparent function of sleep that involves growth process, that of consolidating new long-term memories into neural circuits in the brain.)
The Brain-Maintenance Theory of REM Sleep
If sleep in itself serves purposes of energy conservation, protection from danger, and bodily restoration, then what is the function of REM sleep? Why is restful non-REM sleep interrupted regularly by these periods of increased brain activity and energy expenditure? This question has generated much debate and research.
32
What evidence supports the theories that REM sleep promotes the maintenance of brain circuits?
One longstanding theory is that REM sleep provides regular exercise to groups of neurons in the brain (Hobson, 1988). Synapses can degenerate if they go too long without being active (Edelman, 1987), so neural activity during REM sleep may help preserve important circuits. One line of evidence for this theory is that the longer a person or animal sleeps, the greater is the proportion of sleep time spent in REM sleep. With longer sleep periods there may be more need to interrupt non-REM sleep with exercise.
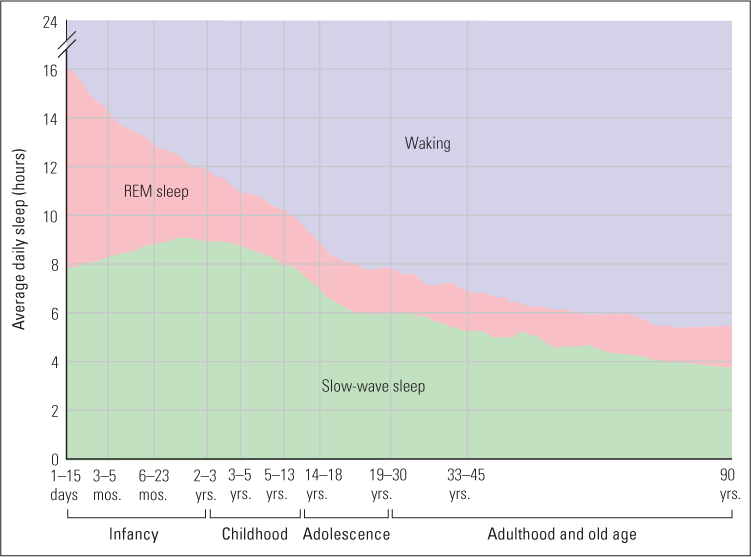
The theory also helps explain why REM sleep occurs to a much greater degree in fetuses and infants than in adults, regardless of species (see Figure 6.12). In fact, the peak of REM sleep in humans occurs in 30-day-old fetuses, which spend almost 24 hours a day in this state. Perhaps as their brains are developing in the relative isolation of the womb, they need to exercise sensory and motor pathways, and REM sleep is their means for doing that (Hobson, 1988). In the fetus, REM sleep is accompanied by body movements such as kicking and twisting, which are apparently triggered by the bursts of activity in motor areas of the brain, so muscles as well as brain circuits are exercised. By the time of birth a neural inhibitory system matures, which inactivates most motor neurons during REM sleep and thus prevents most movements that would otherwise occur. However, the motor neurons to the eyes and to various internal structures, such as the heart, remain uninhibited, so eye movements and increased heart rate persist as observable effects of the brain’s activity.
227
Do Dreams Have Functions?
As just noted, there is good reason to think that REM sleep serves useful functions, but nobody knows if the dreams that accompany such sleep also serve useful functions. One theory, founded on the observation that dreams so often involve fearful content and negative emotions, is that dreams somehow provide a means of rehearsing and resolving threatening experiences that either have happened or could happen in the person’s real life (Valli et al., 2008).
33
How might dreams be explained as inevitable consequences of the state of the brain during REM sleep?
Other sleep researchers, however, have suggested that dreams may not serve any life-promoting functions, but may simply be side effects of the physiological changes that occur during REM sleep (Antrobus, 2000; Hobson, 2002, 2004). Neurons in visual and motor areas of the brain become active during REM sleep, and hallucinations of sights and movements may be an inevitable consequence of such activity. Neurons involved in memory retrieval and emotions also become active, and these may bring familiar images and strong emotional feelings into the sleeping person’s mind. In research done in the 1960s, electrical stimulation in portions of the cerebral cortex produced dream-like hallucinations in people who were awake (Penfield & Perot, 1963). A similar phenomenon may well occur in REM sleep. In addition to producing hallucinations, the brain continues in REM sleep to engage in some degree of thought, just as it does in non-REM sleep. But now the thought becomes wrapped up in trying to make sense of the hallucinations. The result is the weaving of a story connecting one hallucination to the next—hence, the dream. Because of reduced mental capacity during sleep, the story is less logical than one the awake brain would develop, but it still contains some degree of logic.
Sometimes the side-effect theory just described is interpreted as an argument against the psychoanalytic view that dream analysis can be useful for understanding a person’s mind. But that interpretation seems unjustified. Even if dreams are triggered by random events in the brain, the actual images, emotions, and story lines that constitute the dream are not random. They certainly contain elements based on the dreamer’s experience, and because they occur at a time of reduced mental capacity, ideas or feelings that are normally suppressed by higher mental processes could emerge and perhaps be useful in psychoanalysis (Reiser, 1991). In fact, in one experiment, people were more likely to dream about a particular person if they were asked to suppress thoughts about that person just before going to sleep than if they were asked to think actively about that person just before going to sleep (Wegner et al., 2004). This finding is at least consistent with the idea that dream analysis might reveal ideas and concerns that a person is actively suppressing during the day.
228
Individual Variations in the Sleep Drive, and Effects of Failure to Satisfy That Drive
34
How does insomnia differ from nonsomnia? What negative consequences occur when people fail to satisfy their sleep drive?
The sleep drive varies from person to person. Some need more sleep than the typical 8 hours a night to function well, and others need less (Douglas, 2002). At the extreme are rare people, referred to as nonsomniacs, who sleep much less than most of us and yet do not feel tired during the day. A study of such people conducted in the 1970s by Ray Meddis (1977) found that they were generally vigorous and healthy. The most extreme nonsomniac in Meddis’s sample was a 70-year-old nurse who reported that for most of her life she had slept about 50 minutes per night. She was very active during the day and usually spent the night in quiet activities, such as reading or painting. To verify her nonsomnia, Meddis observed her continuously for a prolonged period in the sleep lab. She slept not at all the first 3 days and nights in the lab, remaining cheerful and talkative throughout. Finally, on the fourth night, she slept a total of 99 minutes and awoke feeling fully rested. A more recent study, of people who did well on an average of 3 to 6 hours of sleep per night, found that these people were unusually energetic and scored higher than average, in a positive direction, on an “Attitude to Life” questionnaire (Monk et al., 2001).
The fact that nonsomnia is compatible with physical and psychological health adds to the evidence that only a relatively small amount of sleep is needed for body repair and growth of new synapses in the brain. Yet, most of us do need roughly 8 hours of sleep to function well. We need that sleep because we have a sleep drive that overwhelms our mind and makes us tired, miserable, and relatively ineffective at mental tasks when we fail to meet it. The drive for that much sleep may have evolved for reasons other than body repair and brain growth, but that doesn’t mean we can ignore it.
It is important to distinguish nonsomnia, which is very rare, from insomnia, which is relatively common. An insomniac is someone who has a normal drive for sleep but who, for some reason (such as worry), has great difficulty sleeping at night. Unlike a nonsomniac, an insomniac feels tired during the day as a result of not sleeping. And so do most people who voluntarily reduce their sleep.

Many laboratory studies have been conducted in which people with normal sleep drives voluntarily stay awake for periods of 3 or 4 days or even longer. After about 72 hours awake, some people begin to experience symptoms such as distorted perceptions and extreme irritability (Borbély, 1986). Sleepiness waxes and wanes during such studies, in accordance with the inner clock that controls it. People find it much harder to stay awake during the late night and early morning hours than they do during the rest of the 24-hour day, even after several days of sleep deprivation. In such experiments, scores on tests of vigilance, judgment, and creative thinking also wax and wane in a 24-hour cycle, keeping pace with sleepiness (Horne, 1979, 1988; Jennings et al., 2003). Scores on such tests decline when sleepiness rises, apparently because sleepy people have difficulty attending to the task and because their performance is often interrupted by brief moments of falling asleep, from which they arouse themselves. In general, stimulants such as caffeine, which counteract sleepiness, also remove the negative effects of sleep deprivation on the performance of such tasks. For an example of data showing the effects both of sleepiness and of stimulants on a test of vigilance, see Figure 6.13 (Wesensten et al., 2002).
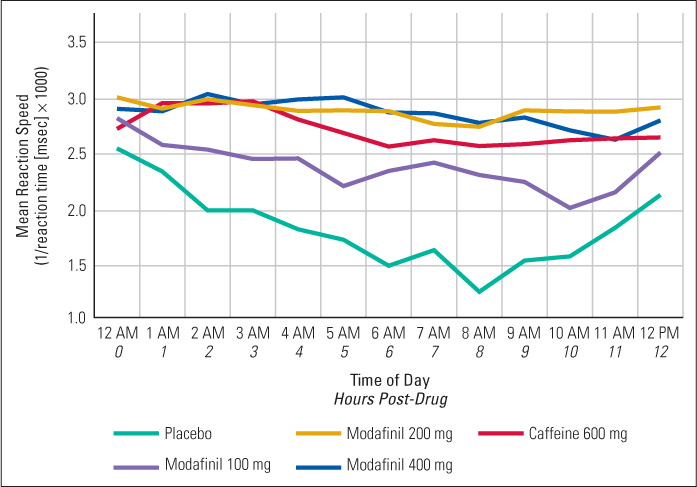
229
In the real world outside of the laboratory, sleepiness—with its accompanying decline in attention and judgment—is dangerous. Many accidents in the workplace result from it, and sleepiness rivals drunkenness as a leading cause of traffic fatalities (Horne & Reyner, 1995, 2001).
Brain Mechanisms Controlling Sleep
In the early years of sleep research, some researchers believed that sleep is the natural state that the brain slips into when not aroused by external stimulation, so they saw no need to posit the existence of special sleep-inducing mechanisms. But such a view is inconsistent with the observation that sometimes sleepiness overwhelms us even when external stimulation is high, while at other times we can’t sleep no matter how quiet, dark, and nonstimulating the environment may be. We now know that sleepiness, like other drives, is actively promoted by neural mechanisms located in the hypothalamus and in brain areas closely connected to the hypothalamus. There are several such neural mechanisms (Pace-Scott & Hobson, 2012), an important one being associated with the daily cycles of sleep and wakefulness, which we describe below.
Rhythm-Generating Neurons in the Hypothalamus Control the Daily Cycle of Sleepiness
35
What is some evidence that the sleep drive is affected by an internal clock, located in the hypothalamus, that can operate even without external time cues?
In all vertebrates, as we noted earlier, the sleep drive waxes and wanes in a cyclic manner over the 24-hour day. This cycle of sleepiness and wakefulness continues, in laboratory animals and human volunteers, even after many days in an artificial “time-free environment”—an environment where there are no regular changes in lighting or other cues that could indicate the time of day. In such an environment, the cycle is typically a few minutes longer or shorter than 24 hours, and it varies from individual to individual, but it is remarkably constant within a given individual (Lavie, 2001).
230
The technical term for any repetitive biological change that continues at close to a 24-hour cycle in the absence of external cues is circadian [sir-kăd′-ē-ǝn] rhythm. The term comes from the Latin words circa, meaning “about,” and dies, meaning “day.” The clock that controls the circadian rhythm of sleep in all mammals is located in a specific nucleus of the hypothalamus called the suprachiasmatic [sup-ra-kī-as-măt′-ic] nucleus (Weaver & Emery, 2012). If this nucleus is damaged, animals lose their regular sleep-wake rhythms and sleep at rather random times over the 24-hour day, and the same is true of human patients (Cohen & Albers, 1991). This nucleus contains rhythm-generating neurons that gradually increase and decrease their rate of action potentials over a cycle of approximately 24 hours, even when surgically isolated from other parts of the brain (Herzog, 2007).
In addition to controlling sleepiness, the suprachiasmatic nucleus also controls a daily rhythm of body temperature (which normally declines in the evening and increases in the morning) and of certain hormones. The hormone that is most directly locked to the circadian clock, and is often used by researchers as an index of the clock’s timing, is melatonin. This hormone, produced by the pineal gland, begins to be secreted into the bloodstream in the evening, typically about 2 hours before a person is ready to fall asleep, and is secreted at relatively high levels until approximately the time when the person is ready to awaken naturally in the morning (Dumont & Beaulieu, 2007).
Input from the Eyes Synchronizes the Hypothalamic Clock to the Light-Dark Cycle
36
What is some evidence that the internal clock is continuously reset by daily changes in light? Through what pathway does that resetting occur?
Under normal conditions, the circadian clock is synchronized with the 24-hour day by the regular waxing and waning of daylight, so rhythms occur in periods of exactly (rather than approximately) 24 hours. Experiments with animals show that the cycle can be lengthened or shortened, by as much as a couple of hours either way, by artificially changing the period of light and dark. Other experiments, with humans as well as nonhuman animals, show that the cycle can be reset through exposure to bright fluorescent lights (Czeisler et al., 1989, 1990).

In general, bright light in the morning and/or dim light or darkness in the evening advances the cycle so that the person becomes sleepy earlier in the evening. In contrast, bright light in the evening and/or avoidance of light in the morning has the opposite effect. It delays the cycle so that sleepiness occurs later at night. A series of several days of such changed conditions can alter the cycle significantly (Dumont & Beaulieu, 2007).
This knowledge about the effects of light has been put to practical use. In one clinical experiment, for example, people with sleep-onset insomnia, inability to fall asleep until very late at night or the wee hours of the morning, were cured or at least partly cured of their insomnia by morning treatments with bright light. After a week of such exposure, the subjects showed an earlier peak of melatonin than before, earlier sleep onset, and an increase of nearly an hour’s more sleep per night, on average, than before treatment. This improvement lasted throughout a 3-week follow-up period after the treatment had ended (Lack et al., 2007). Other research suggests that a tendency to avoid morning light and to use relatively bright lights at home in the evening may be a cause of sleep-onset insomnia in many people today (Dumont & Beauliu, 2007). If you suffer from this problem, you might shift your own cycle to an earlier sleep time by spending some time every morning in sunlight and dimming your room lights in the evening.
231
Brain researchers have found that changes in lighting influence the rhythm-generating neurons by way of a neural tract that runs from the retinas of the eyes to the suprachiasmatic nucleus. These neurons differ from those that are involved in vision, and they derive from light receptors in the retina that differ from the receptors (rods and cones) that are essential for vision (Van Gelder, 2008).
SECTION REVIEW
There is a drive for sleep, a state involving relative unresponsiveness to the environment.
Basic Characteristics of Sleep
- Researchers use EEG records to divide sleep into stages, with stage 4 being the deepest.
- A sleeper progresses through the sleep stages, from light to deep and rapidly back to light, in approximately 90-minute cycles.
- REM (rapid-eye-movement) sleep occurs at the transition between one sleep cycle and the next.
- Most true dreams occur in REM sleep, and sleep thought occurs in non-REM sleep.
Functions of Sleep
- The preservation and protection theory asserts that sleep is needed to conserve energy and increase safety. It is supported by cross-species comparisons of amount and timing of sleep.
- The body-restoration theory contends that the body needs sleep to recover from wear and tear. It is supported by sleep-deprivation studies in animals.
- REM sleep may function to maintain brain circuits and consolidate new learning.
- Different people need different amounts of sleep. Sleep deprivation reduces performance, especially on tests of vigilance.
Brain Mechanisms
- The suprachiasmatic nucleus in the hypothalamus acts as an internal clock for sleepiness and wakefulness.
- Light synchronizes the internal clock with the light-dark cycle.