3.2 How We See and How We Hear
Our two most dominant senses are vision and hearing. This section will first discuss how our eyes and ears gather and process information from the environment. Visual and auditory stimuli are physical energies in the form of waves—
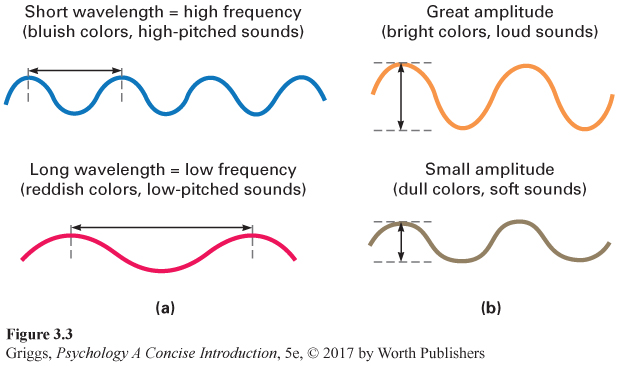
wavelength The distance in one cycle of a wave, from one crest to the next.
amplitude The amount of energy in a wave, its intensity, which is the height of the wave at its crest.
Wavelength refers to the distance in one cycle of a wave, from one crest to the next. Different wavelengths of light lead to varying perceptions of color. Humans can perceive wavelengths roughly from 400 to 700 nanometers (nm), or billionths of a meter. As the wavelengths decrease from 700 nm, the resulting hues go from red through the colors of the spectrum down to violet. One way to remember the order of these colors is by remembering the name ROY G. BIV, which stands for Red, Orange, Yellow, Green, Blue, Indigo, and Violet. Amplitude refers to the amount of energy in a wave, its intensity, which is the height of the wave at its crest. For light waves, the amplitude determines the brightness of the light. Different amplitudes lead to different levels of brightness; the greater the amplitude, the brighter the light.
frequency The number of times a wave cycles in one second.
For auditory stimuli, the frequency of the waveform rather than its length is used. Frequency refers to the number of times the waveform cycles in one second. Longer wavelengths have lower frequencies, and shorter wavelengths have higher frequencies. The frequency of a sound wave determines its pitch, the quality of sound that we describe as high or low. For example, female voices usually have a higher pitch than male voices. Humans can perceive sound wave frequencies from about 20 to 20,000 hertz (Hz), the number of cycles of the sound wave in one second. This unit of measurement’s name, hertz, comes from a nineteenth-
transduction The conversion of physical energy into neural signals that the brain can understand.
The brain cannot process physical energies such as light waves and sound waves. There are receptor cells in our eyes and ears that specialize in transduction—converting physical energy into neural signals that the brain can understand. This conversion is the first step in processing incoming sensory information. Following transduction, the incoming visual and auditory signals undergo further processing on their way to the brain, and within the brain before we see and hear the brain’s interpretations of these signals. Next, we will take a closer look at transduction and the initial information processing in the eyes and ears, focusing on how we see color and how we distinguish the pitches of sounds.
How the Eye Works
The light-
accommodation The focusing of light waves from objects of different distances directly on the retina.
The light then passes through the transparent lens, which is responsible for accommodation—the focusing of the light waves from objects of different distances directly on the retina. This process is called accommodation because the lens changes its shape (accommodates) in order to focus objects at different distances. When looking at distant objects (at least 20 feet away), the lens is in its unaccommodated flattest state, which allows the light waves to be focused on the retina. When we look at objects that are nearer, the lens accommodates by becoming thicker in order to focus the light waves properly on the retina. Because light travels in straight lines, the retinal image will be inverted and reversed. The brain, however, rearranges this image to its correct orientation for our perception.
nearsightedness A visual problem in which the light waves from distant objects come into focus in front of the retina, blurring the images of these objects.
farsightedness A visual problem in which the light waves from nearby objects come into focus behind the retina, blurring the images of these objects.
If the image is not properly focused on the retina because of defects in the lens or the shape of the eyeball, problems in vision arise. Nearsightedness occurs when the light waves from distant objects come into focus in front of the retina, which blurs the images of these objects. Farsightedness is created by the opposite focusing problem—
It is also important to realize that our brains do not process everything in the physical world but rather only a tiny bit of it. For example, the part of the light spectrum that is visible to us is less than a ten-
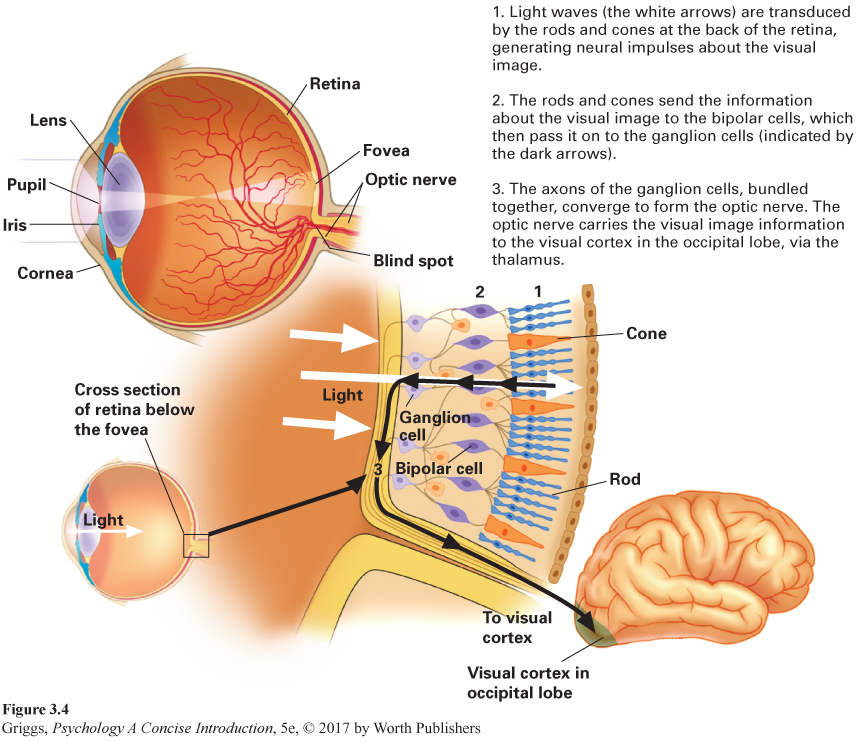
retina The light-
The structure of the retina. Except for the visual image being reversed and inverted, no visual processing occurs until the light waves reach the retina, which is only a mere half-

The optic nerve carries the visual information to the thalamus, which is responsible for directing it to the primary visual cortex in our occipital lobes to be processed. But how does the visual cortex interpret this message from the retina? David Hubel and Torsten Wiesel’s innovative research in which they recorded patterns of electrical impulses in individual neurons in the visual cortex of cats provided the answer. They were awarded the Nobel Prize in Physiology or Medicine in 1981 for their discoveries concerning information processing in the visual system. The story of their collaboration along with reprints of all of their key publications can be found in Hubel and Wiesel’s Brain and Visual Perception: The Story of a 25-

Essentially, the brain recognizes a visual object (a process called pattern recognition) by breaking it down into its elementary features, such as angles and diagonal lines. Cortical cells called feature detectors then recognize these elementary features. Finally, the elementary features are put back together again, so that the whole object can be recognized. For example, to recognize a triangle, Δ, it is broken down into its features (possibly, /, ̲ , and \), which are recognized and then regrouped into the proper relationship. Strange isn’t it? To recognize an object, the object is broken down into its parts and then these parts are put back together again in order to identify object. Even stranger is that there are separate processing systems for form, color, distance, and motion information, which the brain somehow coordinates and unifies into our seamless perception of the world (Ratey, 2001).
blindsight A condition in which a blind person has some spared visual capacities in the absence of any visual awareness.
But what if your primary visual cortex were severely damaged? You would be blind in the conventional sense, but you might possibly have what is called blindsight (seeing without knowing)—a condition in which a blind person has some spared visual capacities in the absence of any visual awareness (Weiskrantz, 2009). The oxymoron blindsight was coined by Lawrence Weiskrantz of Oxford University, who has studied this condition for more than 40 years (Weiskrantz, Warrington, Sanders, & Marshall, 1974). People with blindsight respond to a visual stimulus without consciously experiencing it. For example, they claim that they do not see an object, but when asked to reach out and touch it, they can do so far more often than what chance would predict. If you put a large object in their path, they will walk around it but insist that they do not see anything. The experimental evidence concerning blindsight has sometimes been controversial (Cowey, 2010), and the explanation for blindsight remains unsettled. Some researchers believe that the blindsight responses stem from the flow of visual information through neural pathways that bypass the damaged visual cortex (a kind of information detour) to brain areas that do not convey the conscious perception but enable the blindsight responses (Ptito & Leh, 2007; Schmid et al., 2010). An alternative explanation is that a small amount of healthy tissue remains within the damaged visual cortex, not enough to provide conscious perception but sufficient for blindsight responses (Gazzaniga, Fendrich, & Wesainger, 1994). It is also possible that both hypotheses are correct. Because there is much variability between those with blindsight, each hypothesis might explain different cases. One conclusion that can be made, however, is that normal visual perception requires a healthy visual cortex.
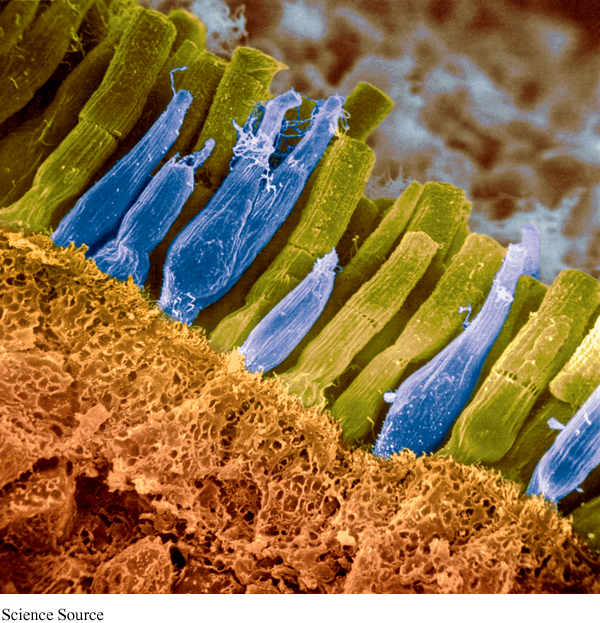
rods Receptor cells in the retina that are principally responsible for dim light and achromatic vision.
cones Receptor cells in the retina that are principally responsible for bright light and color vision.
fovea A tiny pit in the center of the retina filled with cones.
Rods and cones. There are two types of visual receptor cells, rods and cones, in the retina. They are called rods and cones because of their actual physical shapes. Their functions are also very different. Rods are principally responsible for dim light and achromatic (colorless) vision, and cones for bright light and color vision. Rods outnumber cones about 20 to 1 (Kalat, 2007). There are about 120 million rods and only 6 million or so cones in each eye. Cones are more centrally located in the retina than rods. They are located in the fovea, a tiny pit in the center of the retina, and in the periphery of the retina on each side of the fovea. Rods are only located in the periphery, and the proportion of rods to cones increases with increasing distance from the fovea. The major differences between rods and cones are summarized in Table 3.3.
| Rods | Cones |
|---|---|
| 120 million in each eye | 6 million in each eye |
| Located mainly in periphery of retina | Located mainly in fovea and central retina |
| Primarily responsible for dim light vision | Primarily responsible for bright light vision |
| Responsible for achromatic (colorless) vision | Responsible for chromatic (color) vision |
| Lead to low visual acuity | Lead to high visual acuity |
The difference in location of rods (peripheral) and cones (central) helps us to determine where we should focus an object for the best visual acuity (resolution of detail). In normal or brighter light when the cones are mainly responsible for our vision, the object should be focused on the fovea, which is packed with cones. Cones provide a clearer picture of an object. Why? Cones, especially foveal cones, tend to have more direct routes than the rods to the bipolar and ganglion cells for the retinal information that they process. For example, typically one foveal cone communicates with one bipolar cell, but several rods communicate with one bipolar cell. This means that the information that the rods send forward has been averaged across several retinal positions, leading to a less clear picture of the retinal image. Foveal cones do not have to do such averaging; therefore, the retinal image they send forward is more detailed and clearer. The cones in the periphery just outside of the fovea do some averaging but far less than the rods. A good example of the foveal cones’ role in the resolution of detail concerns the changes in French artist Edgar Degas’s paintings as his retinal disease worsened in his later years. As his central vision grew blurry, his paintings became less detailed and coarser and lost refinement (Marmor, 2006). He could not see the detail so he could not paint it.
dark adaptation The process by which the rods and cones through internal chemical changes become more and more sensitive to light in dim light conditions.
But where should we focus an object in dim light conditions? The answer is in the periphery of the retina rather than the fovea. Why? This is where the rods are more densely located, and they are mainly responsible for our vision when not much light is available. We shouldn’t look directly at the object when there isn’t much light, but rather focus a little to the side of it so that it will be projected onto the periphery of the retina. The image won’t be as clear as when we can use our cones, but it is the best we can do in dim light. The major role of the rods in dim light vision is also apparent in dark adaptation, the process by which the rods and cones through internal chemical changes become more and more sensitive to light in dim light conditions. The cones adapt quickly (in 5 to 10 minutes), but need more light to function, so they do not really help us to see in dim light. It takes longer for the rods to adapt (20 minutes or so), but they slowly help us to see in dim light. Normally, however, we have enough light for our cones to be working. That is why we see the world in vivid color. But how do the cones do this, and what else is involved in producing our perception of color? These are the questions we discuss next.
How We See Color
How many different colors can humans discriminate? Hundreds? Thousands? These estimates are not even close. It has been estimated that the human eye can discriminate about 7.5 million different colors (Fineman, 1996). How can we differentiate so many colors? Actually our color vision can be explained by combining two rather simple theories of color vision, the trichromatic theory and the opponent-
trichromatic theory A theory of color vision that assumes that there are three types of cones, each only activated by wavelength ranges of light corresponding roughly to blue, green, and red. It further assumes that all of the various colors that we can see are mixtures of various levels of activation of the three types of cones. If all three are equally activated, we see white.
Trichromatic theory of color vision. Trichromatic theory, as the word trichromatic implies, proposes that there are three colors involved. It assumes that there are three types of cones, activated best by short, medium, or long wavelengths of light, roughly corresponding to blue, green, and red. Indeed, physiological research indicates that there are three types of cones, each containing a slightly different photopigment that is maximally sensitive to blue, green, or red wavelengths of light (Wald, 1964). The trichromatic theory assumes that all of the many colors that we can perceive are mixtures of various proportions of these three cone activities. If all three types of cones are equally active, we see white (the sum of the three wavelengths of light). Trichromatic theory played a role in the creation of the television screen, which consists of microscopic red, green, and blue dots. The colors we see on the screen are the product of how these three types of dots are activated by the television broadcast and how the light-
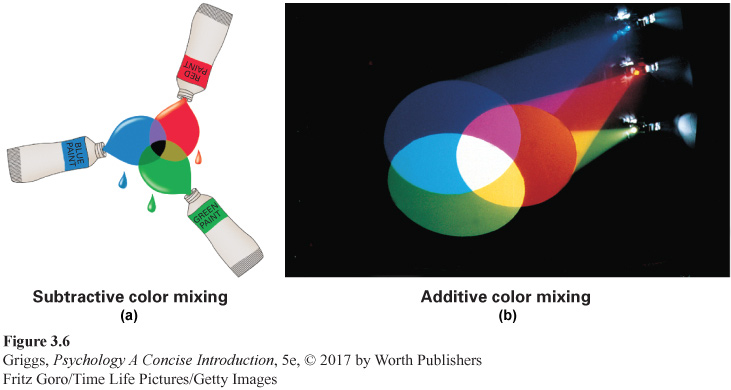
additive mixtures Direct mixtures of different wavelengths of light in which all of the wavelengths reach the retina and are added together.
subtractive mixtures Mixtures of wavelengths of light in which some wavelengths are absorbed (subtracted) and so do not get reflected from the mixtures to the retina.
It is important to realize that the proposed mixtures of the primary colors are additive mixtures—different wavelengths of light are directly mixed together. In additive mixtures, all of the wavelengths of light reach the retina and are added together. The resulting colors are very different from those for subtractive mixtures, such as mixing paints. In subtractive mixtures, some wavelengths are absorbed (subtracted) and so do not get reflected from the mixture to the retina. For example, if we mix red, green, and blue paints together in equal amounts, the result is black (no light reflected). A mixture of equal proportions of red, green, and blue wavelengths of light, however, appears white. These subtractive and additive mixtures are shown in Figure 3.6.
The trichromatic theory provides a good explanation of the most prevalent type of color blindness—
complementary colors Wavelengths of light that when added together produce white.
There are, however, color phenomena that trichromatic theory has difficulty explaining. We know that red-
opponent-
Opponent-
Opponent-

There is research that supports both trichromatic theory and opponent-
| Theory | Explanation of Color Vision |
|---|---|
| Trichromatic theory | There are three types of cones, each only activated by wavelength ranges of light corresponding roughly to blue, green, and red; all colors are mixtures of various levels of activation of these three types of cones |
| Opponent- |
There are three opponent- |
| Composite theory | Color information is processed by the cones according to trichromatic theory, but color information is processed at the post- |
This brief discussion of how we see color shows how visual input from the environment is processed, analyzed, and reconfigured, starting with its initial processing in the retina and as it continues along the visual pathways to the brain to be interpreted. Next, we will consider how such processing of auditory information enables us to hear.
How the Ear Works
The auditory sensory system is a mechanical (vibratory) system with the receptor cells located in the inner portion of each ear. As you can see in Figure 3.8, the ear is divided into three sections—
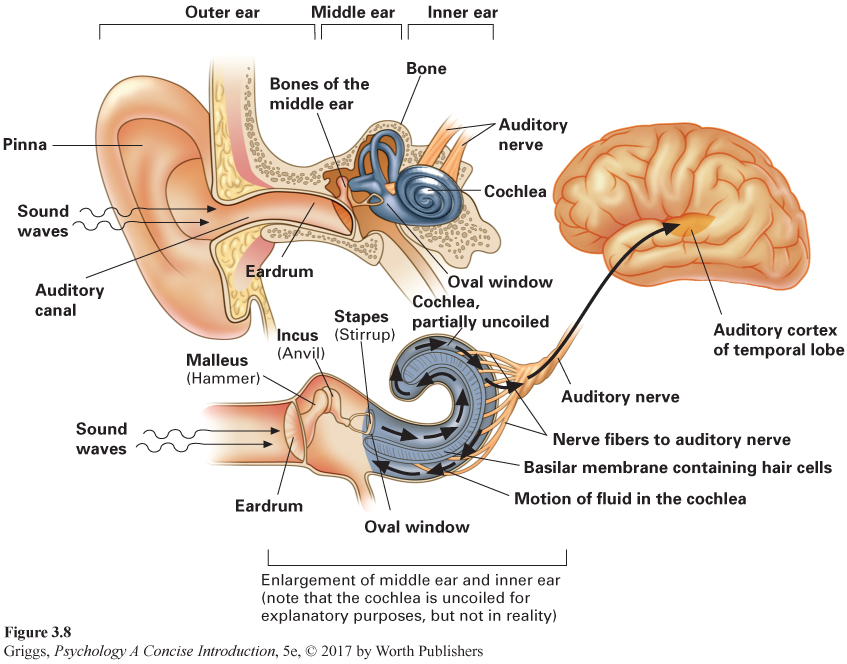
cochlea A snail-
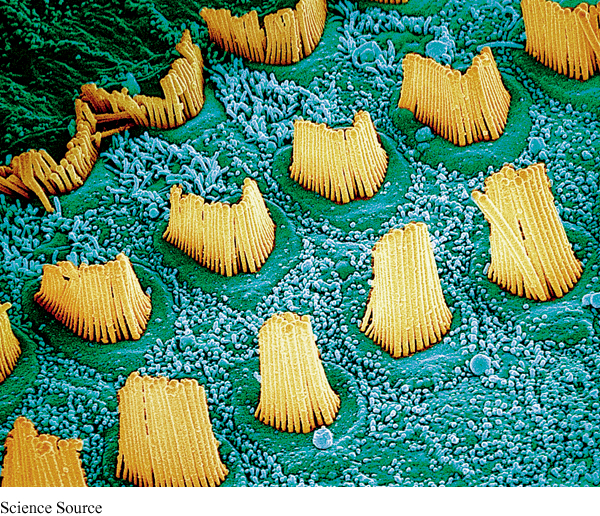
hair cells The receptor cells for hearing. They line the basilar membrane inside the cochlea.
The cochlea is a snail-
nerve deafness Hearing loss created by damage to the hair cells or the auditory nerve fibers in the inner ear.
conduction deafness Hearing loss created by damage to one of the structures in the ear responsible for mechanically conducting the auditory information to the inner ear.
Hearing loss caused by damage to these hair cells or the auditory nerve fibers is called nerve deafness. The most common causes of such damage are aging and continued exposure to extremely loud noises. Nerve deafness is much more difficult to treat than conduction deafness, hearing loss caused by damage to the mechanical system carrying the auditory information to the cochlea. For example, puncturing an eardrum can lead to conduction deafness. Hearing aids can often help to alleviate this type of deafness.
It is important to realize that just as we learned that we only process a small part of the light spectrum, parts of the auditory spectrum are outside the range of human hearing. We cannot process ultrasound stimuli, those above 20,000 Hz, and infrasound stimuli, those below 20 Hz. Other animals, however, have different windows on the auditory world. For example, elephants, whales, and giraffes can hear infrasound stimuli; and dolphins and bats, ultrasound stimuli. Whales and elephants use infrasound to communicate over long distances, possibly up to hundreds of miles in the case of whales. Dolphins communicate using ultrasound frequencies, and bats use them to navigate and hunt in darkness. Dogs and cats can hear frequencies above 40,000 and 50,000 Hz, respectively; and the upper range for dolphins may extend to 150,000 Hz (Goldstein, 2007). Thus, the outside world is really noisy; we just lack the ability to process it.
Now, given our basic understanding of how the ear works and how auditory information gets coded and processed, we’ll consider an important aspect of this complex coding, how we distinguish different pitches.
How We Distinguish Pitch
Remember that pitch is the quality of sound that we describe as high or low and is determined by the frequency of a sound wave. We can perceive sound wave frequencies from about 20 to 20,000 Hz. Each of the frequencies in this rather large range corresponds to a different pitch that we can perceive. The best explanation of how we perceive this range of pitches parallels the story of how we see different colors. We will find that to get the best explanation, two theories will need to be combined—
place theory A theory of pitch perception that assumes that there is a specific location along the basilar membrane that will maximally respond to a particular frequency, thereby indicating the pitch to the brain. As this location goes down the basilar membrane from the oval window, the pitch goes down from 20,000 Hz to 20 Hz.
Place theory. Place theory assumes that there is a specific place along the basilar membrane, starting from the oval window, which will respond maximally to a particular frequency. Thus, the brain will know the frequency by knowing the location of peak activity along the basilar membrane. In some ingenious experiments on the cochleas from human cadavers, Georg von Békésy discovered that each frequency generates a traveling wave that moves along the basilar membrane and peaks at a particular location (Békésy, 1960). Starting at the oval window and 20,000 Hz, the pitch goes down as the peak activity moves down the basilar membrane. These peak locations tell the brain the frequencies of the incoming auditory stimuli.
frequency theory A theory of pitch perception that assumes that the frequency of the sound wave is mimicked by the firing rate of the entire basilar membrane.
Frequency theory. Frequency theory assumes that the frequency of the sound wave is mimicked by the firing rate of the hair cells across the entire basilar membrane. For example, if the frequency were 100 Hz, the firing rate for the membrane would be 100 impulses per second. The brain would know the frequency by the basilar membrane’s firing rate in reaction to the auditory stimulus.
volley principle Cells taking turns firing will increase the maximum firing rate for a group of cells.
Now let’s see why and how we put theories together to get a better explanation of pitch perception. First, research has found that specific locations along the basilar membrane correlate well with the frequencies of sound waves, except for lower pitches (< 500 Hz). For these lower pitches, the firing rate of the basilar membrane mimics the frequency. This means that place theory can explain how we hear the entire range of pitches except for low pitches, but frequency theory can explain how we hear these low pitches. Is frequency theory only able to explain how we perceive pitches below 500 Hz? No, the upper limit on firing rate is 5,000 times per second. There is a maximum firing rate for nerve cells, about 1,000 times per second. However, with the addition of the volley principle, in which the cells take turns firing to increase the maximum firing rate for the group of cells, frequency theory could explain our perception of frequencies up to about 5,000 Hz (Zwislocki, 1981). Because 5,000 times per second is the upper limit for the firing rate using the volley principle, frequency theory would not be able to explain how we perceive higher frequencies of 5,000 to 20,000 Hz.
Combining the two theories gives us a composite theory that fits the research findings on pitch perception. Frequency theory explains the perception of lower frequencies (< 500 Hz), and place theory explains how we perceive higher frequencies (> 5,000 Hz). This means that the brain uses the firing rate to differentiate the low frequencies and the location of maximal hair cell activity along the basilar membrane to distinguish the high frequencies. For the range of frequencies in between (roughly 500 to 5,000 Hz), both sources of information work, giving us double coverage and better pitch perception. This is fortunate because this range of frequencies contains the sounds most important to us—
| Theory | Explanation of Pitch Perception |
|---|---|
| Place theory | There is a specific location along the basilar membrane that will maximally respond to a particular frequency, thereby indicating the pitch to the brain |
| Frequency theory | The frequency of the sound wave is mimicked by the firing rate of the entire basilar membrane, thereby indicating the pitch to the brain |
| Composite theory | Lower frequencies (< 500 Hz) are processed according to frequency theory, higher frequencies (> 5,000 Hz) are processed according to place theory, and for frequencies between from 500 to 5,000 Hz, both theories are working |
Section Summary
In this section, we discussed our two most dominant senses—
Light waves pass through many parts of the eye before being transduced by the rods and cones at the back of the retina. Once transduced, the neural signals are carried by the optic nerve to the thalamus, which sends them to the primary visual cortex where they are interpreted. In this interpretation process, the object is broken down into its elemental features, which are recognized by feature detectors, and then the features are combined so that the whole object can be recognized. The cones and rods divide the initial visual processing. Cones are responsible for bright light and color vision; rods are responsible for dim light and achromatic vision.
There are two competing explanations for how we see color—
The auditory system is a mechanical vibratory system with the hair cell receptors located within the basilar membrane in the cochlea. Incoming vibrations result in fluid displacement in the cochlea that causes movement of these hair cells. The nature of these hair cell movements creates the auditory message that gets carried by the auditory nerve, by way of the thalamus, to the primary auditory cortex for interpretation. Damage to these hair cells or the auditory nerve results in nerve deafness, which is very difficult to treat. Damage to one of the structures conducting the auditory information to the inner ear causes conduction deafness, which is easier to treat.
To explain how we perceive pitch, two theories are necessary. We can perceive a fairly large range of frequencies, 20 to 20,000 Hz. Place theory, which assumes that there are specific locations along the basilar membrane correlated with each frequency, explains how we hear high pitches (frequencies above 5,000 Hz). Frequency theory, which assumes that the firing rate of the hair cells mimics the incoming frequency information, explains how we hear low pitches (frequencies less than 500 Hz). Frequency theory (with the addition of the volley principle) and place theory both operate to give us better pitch perception for moderate pitches (frequencies from 500 to 5,000 Hz).
2
Question 3.5
.
Explain the difference between nearsightedness and farsightedness with respect to focusing problems.
In nearsightedness, we have difficulty viewing distant objects because their images come into focus in front of the retina; in farsightedness, we have difficulty viewing near objects because their images come into focus behind the retina. The focusing problems could be due to defects in the lens or the shape of the eyeball.
Question 3.6
.
Explain what opponent-
After staring at the flag with alternating yellow and white stripes and a block of green in the middle, the yellow, white, and green parts of the three opponent-
Question 3.7
.
Explain why longer sound waves lead to lower frequencies and shorter ones lead to higher frequencies.
Longer wavelengths lead to lower frequencies because such wavelengths can only cycle a small number of times per second. Similarly, shorter wavelengths lead to higher frequencies because they can cycle more times per second.
Question 3.8
.
Explain why neither the place theory nor the frequency theory by itself can explain human perception of pitch from 20 to 20,000 Hz.
Neither theory by itself can explain how we hear the entire range of pitches, 20 to 20,000 Hz, because each theory is unable to explain pitch perception for a particular part of this range. Place theory cannot explain how we perceive low pitches (< 500 Hz) because there are no places of maximal firing along the basilar membrane for these frequencies. The firing rate of the entire membrane mimics these frequencies. Similarly, frequency theory cannot explain how we perceive high pitches, those greater than 5,000 Hz, because there is a physiological limit on the firing rate for cells. Even if the volley principle is employed, this limit is about 5,000 times per second. This means that the hair cells could not generate firing rates to match the high frequencies.