CONCEPT20.1 Eukaryotes Acquired Features from Both Archaea and Bacteria
We easily recognize trees, mushrooms, and insects as plants, fungi, and animals, respectively. But there is a dazzling assortment of other eukaryotic organisms—mostly microscopic—that do not fit into these three groups. Eukaryotes that are not plants, animals, or fungi have traditionally been called protists. But phylogenetic analyses reveal that many of the groups we commonly refer to as protists are not, in fact, closely related. Thus the term “protist” does not describe a formal taxonomic group, but is a convenience term for “all the eukaryotes that are not plants, animals, or fungi.”
The unique characteristics of the eukaryotic cell lead scientists to conclude that the eukaryotes are monophyletic, and that a single eukaryotic ancestor diversified into the many different protist lineages as well as giving rise to the plants, fungi, and animals. As we saw in Concept 19.1, eukaryotes are generally thought to be more closely related to archaea than to bacteria. The mitochondria and chloroplasts of eukaryotes, however, are clearly derived from bacterial lineages (see Figure 19.1).
Traditionally, biologists have hypothesized that the split of Eukarya from Archaea was followed by the endosymbioses with bacterial lineages that led to the origin of mitochondria and chloroplasts. Some biologists prefer to view the origin of eukaryotes as the fusion of lineages from the two prokaryote groups. This difference is largely a semantic one that hinges on the subjective point at which we deem the eukaryote lineage to have become definitively “eukaryotic.” In either case, we can make some reasonable inferences about the events that led to the evolution of a new cell type, bearing in mind that the environment underwent an enormous change—from low to high availability of free atmospheric oxygen—during the course of these events.
LINK
You can compare eukaryotic and prokaryotic cells by reviewing Concepts 4.2 and 4.3
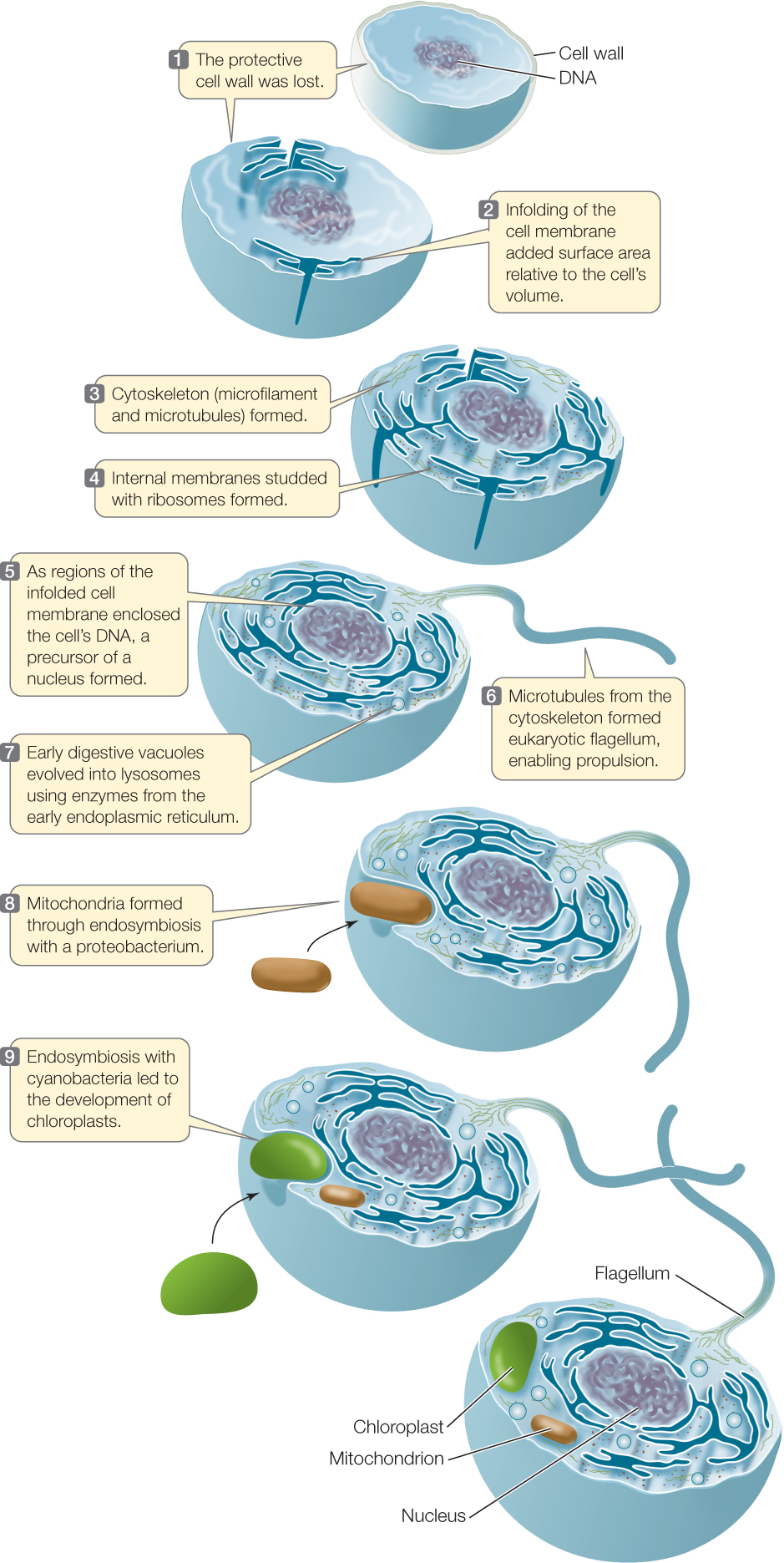
The modern eukaryotic cell arose in several steps
Several events were important in the origin of the modern eukaryotic cell (FIGURE 20.1):
- The origin of a flexible cell surface
- The origin of a cytoskeleton
- The origin of a nuclear envelope, which enclosed a genome organized into chromosomes
- The appearance of digestive vacuoles
- The acquisition of certain organelles via endosymbiosis
Flexible Cell Surface
We presume that ancient prokaryotic organisms, like most present-day prokaryotic cells, had firm cell walls. The first step toward the eukaryotic condition was the loss of the cell wall by an ancestral prokaryotic cell. This wall-less condition occurs in some present-day prokaryotes.
Consider the possibilities open to a flexible cell without a firm cell wall, starting with cell size. As a cell grows larger, its surface area-to-volume ratio decreases (see Figure 4.2). Unless the surface area can be increased, the cell volume will reach an upper limit. If the cell’s surface is flexible, however, it can fold inward and become more elaborate, creating more surface area for gas and nutrient exchange. With a surface flexible enough to allow infolding, the cell can exchange materials with its environment rapidly enough to sustain a larger volume and more rapid metabolism (see Figure 20.1, steps 1–2). Furthermore, a flexible surface can pinch off bits of the environment, bringing them into the cell by endocytosis. These infoldings of the cell surface, which also exist in some modern prokaryotes, were important for the evolution of large eukaryotic cells.
Changes in Cell Structure and Function
Other early steps that were important for the evolution of the eukaryotic cell involved increased compartmentalization and complexity of the cell (see Figure 20.1, steps 3–7):
- The development of a more complex cytoskeleton
- The formation of ribosome-studded internal membranes, some of which surrounded the DNA
- The enclosure of the cell’s DNA in a nucleus
- The formation of a flagellum from microtubules of the cytoskelton
- The evolution of digestive vacuoles
Until a few years ago, biologists thought that cytoskeletons were restricted to eukaryotes. Improved imaging technology and molecular analyses have now revealed homologs of many cytoskeletal proteins in prokaryotes, so simple cytoskeletons evolved before the origin of eukaryotes. The cytoskeleton of a eukaryote, however, is much more developed and complex than that of a prokaryote. This greater development of microfilaments and microtubules supports the eukaryotic cell and allows it to manage changes in shape, to distribute daughter chromosomes, and to move materials from one part of its larger cell to other parts. In addition, the presence of microtubules in the cytoskeleton allowed some cells to develop the characteristic eukaryotic flagellum.
The DNA of a prokaryotic cell is attached to a site on its cell membrane. If that region of the cell membrane were to fold into the cell, the first step would be taken toward the evolution of a nucleus, a primary feature of the eukaryotic cell. The nuclear envelope appeared early in the eukaryote lineage. The next step was probably phagocytosis—the ability to engulf and digest other cells.
Eukaryotes acquired features from both archaea and bacteria
Ribosomal RNA (rRNA) genes are present in the nuclear genome of eukaryotes. There are also rRNA genes in the genomes of mitochondria and chloroplasts. Therefore, photosynthetic eukaryotes have three different sets of rRNA genes, which encode the structural RNA of separate ribosomes in the nucleus, mitochondria, and chloroplasts, respectively. Translation of each genome takes place on its own set of ribosomes.
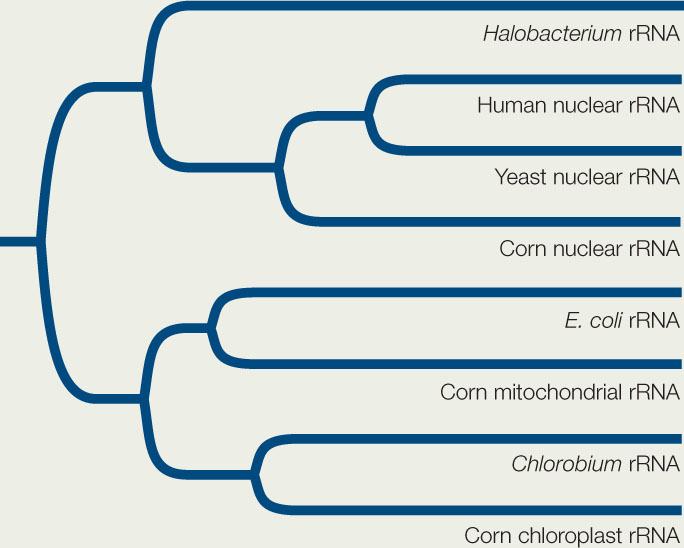
The gene tree shows the evolutionary relationships of rRNA gene sequences isolated from the nuclear genomes of humans, yeast, and corn; from an archaeon (Halobacterium), a proteobacterium (E. coli), and a cyanobacterium (Chlorobium); and from the mitochondrial and chloroplast genomes of corn. Use the gene tree to answer the following questions.
- Why aren’t the three rRNA genes of corn one another’s closest relatives?
- How would you explain the closer relationship of the mitochondrial rRNA gene of corn to the rRNA gene of E. coli than to the nuclear rRNA genes of other eukaryotes? Can you explain the relationship of the rRNA gene from the chloroplast of corn to the rRNA gene of the cyanobacterium?
- If you were to sequence the rRNA genes from human and yeast mitochondrial genomes, where would you expect these two sequences to fit on the gene tree?
Endosymbiosis
At the same time the processes outlined above were taking place, cyanobacteria were generating O2 as a product of photosynthesis. The increasing concentrations of O2 in the oceans, and eventually in the atmosphere, had disastrous consequences for most organisms of the time, which were unable to tolerate the newly oxidizing environment. But some prokaryotes evolved strategies to use the increasing oxygen, and—fortunately for us—so did some of the new phagocytic eukaryotes.
At about this time, endosymbioses began to play a role in eukaryote evolution (see Figure 20.1, steps 8–9). The theory of endosymbiosis proposes that certain organelles are the descendants of prokaryotes engulfed, but not digested, by ancient eukaryotic cells. One crucial event in the history of eukaryotes was the incorporation of a proteobacterium that evolved into the mitochondrion. Initially the new organelle’s primary function was probably to detoxify O2 by reducing it to water. Later this reduction became coupled with the formation of ATP in cellular respiration. Upon completion of this step, the essential modern eukaryotic cell was complete.
LINK
You may wish to review the reactions of cellular respiration in Concept 6.2
Photosynthetic eukaryotes are the result of yet another endosymbiotic step: the incorporation of a prokaryote related to today’s cyanobacteria. This endosymbiont evolved into the modern chloroplast.
Chloroplasts have been transferred among eukaryotes several times
Eukaryotes in several different groups possess chloroplasts, and groups with chloroplasts appear in several distantly related eukaryote clades. Some of these groups differ in the photosynthetic pigments their chloroplasts contain. And not all chloroplasts are limited to a pair of surrounding membranes—in some photosynthetic eukaryotes, they are surrounded by three or more membranes. We now view these observations as evidence of a remarkable series of endosymbioses. This conclusion is supported by extensive evidence from electron microscopy and nucleic acid sequence comparisons.
All chloroplasts trace their ancestry back to the engulfment of one cyanobacterium by a larger eukaryotic cell. This event, the step that first gave rise to the photosynthetic eukaryotes, is known as primary endosymbiosis (FIGURE 20.2A). The cyanobacterium, a Gram-negative bacterium, had both an inner and an outer membrane (see Figure 19.2B). Thus the original chloroplasts had two surrounding membranes: the inner and outer membranes of the cyanobacterium. Remnants of the peptidoglycan-containing cell wall of the bacterium are present in the form of a bit of peptidoglycan between the chloroplast membranes of glaucophytes, the first eukaryote group to branch off following primary endosymbiosis (as we will see in Chapter 21). Primary endosymbiosis also gave rise to the chloroplasts of the red algae, green algae, and land plants. The red algal chloroplast retains certain pigments of the original cyanobacterial endosymbiont that are absent in green algal chloroplasts.
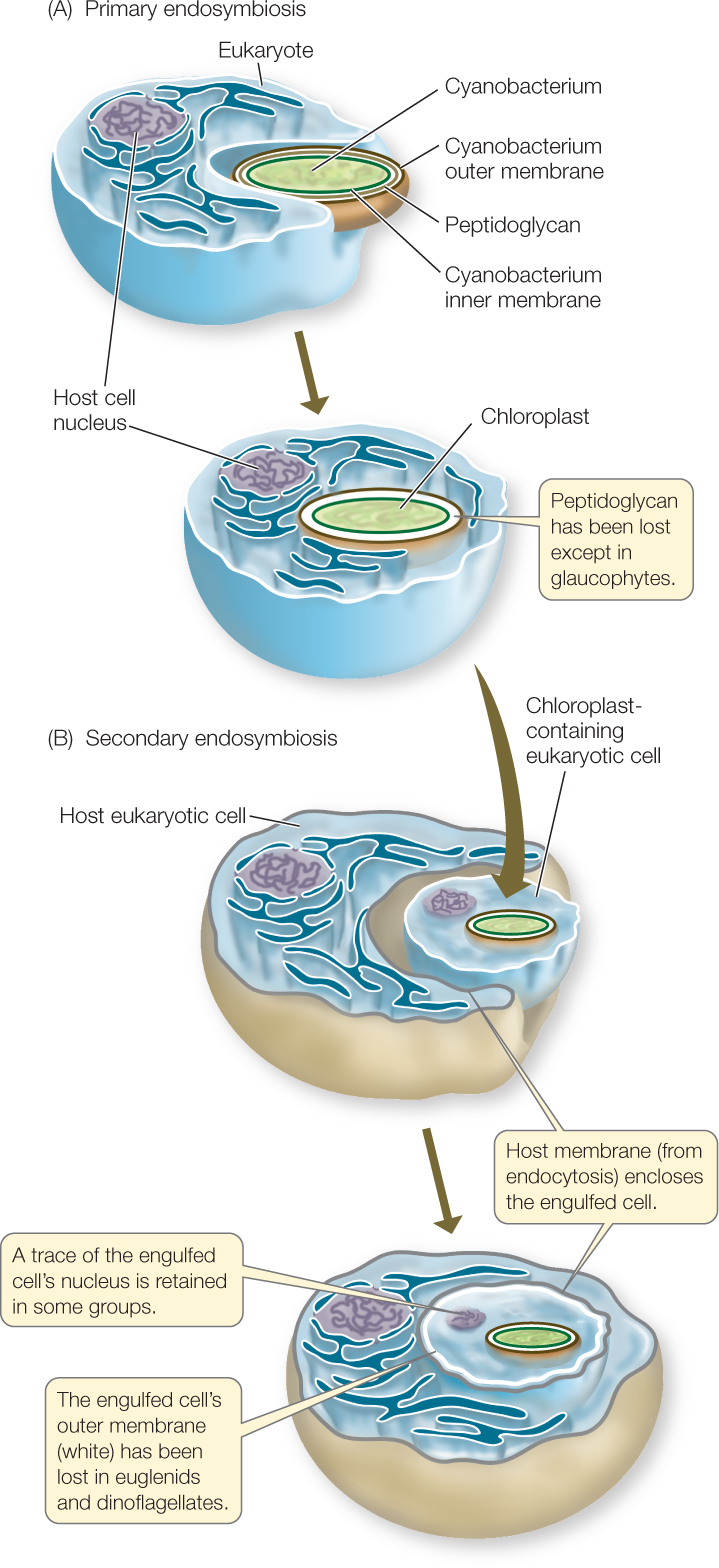
Almost all remaining photosynthetic eukaryotes are the result of additional rounds of endosymbiosis. For example, the photosynthetic euglenids derived their chloroplasts from secondary endosymbiosis (FIGURE 20.2B). Their ancestor took up a unicellular green alga, retaining its chloroplast and eventually losing the rest of the constituents of the alga. This history explains why the photosynthetic euglenids have the same photosynthetic pigments as the green algae and land plants. It also accounts for the third membrane of the euglenid chloroplast, which is derived from the euglenid’s cell membrane (as a result of endocytosis). An additional round—tertiary endosymbiosis—occurred when a dinoflagellate apparently lost its chloroplast and took up another protist that had acquired its chloroplast through secondary endosymbiosis.
CHECKpointCONCEPT20.1
- Why was the development of a flexible cell surface a key event for eukaryote evolution?
- What do you consider the most critical events that led to the evolution of the eukaryotic cell? Why?
- Explain how increased availability of atmospheric oxygen (O2) could have affected the evolution of the eukaryotic cell.
The features that eukaryotes gained from archaea and bacteria have allowed them to exploit many different environments. This led to the evolution of great diversity among eukaryotes, beginning with a radiation that started in the Precambrian.