Concept 23.6: Life on Land Contributed to Vertebrate Diversification
The evolution of lunglike sacs in fishes set the stage for the invasion of the land. Some early ray-finned fishes probably used those sacs to supplement their gills when oxygen levels in the water were low, as lungfishes and many groups of ray-finned fishes do today. But with their unjointed fins, those fishes could only flop around on land. Changes in the structure of the fins first allowed some fishes to support themselves better in shallow water and, later, to move better on land.
Jointed fins enhanced support for fishes
In the lobe-limbed vertebrates, the paired pelvic and pectoral fins developed into more muscular fins that were joined to the body by a single enlarged bone. The modern representatives of these lobe-limbed vertebrates include the coelacanths, lungfishes, and tetrapods.
The coelacanths flourished from the Devonian until about 65 million years ago, when they were thought to have become extinct. However, in 1938 a commercial fisherman caught a living coelacanth off South Africa. Since that time, hundreds of individuals of this extraordinary fish, Latimeria chalumnae, have been collected. A second species, L. menadoensis, was discovered in 1998 off the Indonesian island of Sulawesi. Latimeria, a predator of other fishes, reaches a length of about 1.8 meters and weighs up to 82 kilograms (FIGURE 23.43A). Its skeleton is composed mostly of cartilage, not bone. The cartilaginous skeleton is a derived feature in this clade because it had bony ancestors.


Go to MEDIA CLIP 23.14 Coelacanths in the Deep Sea
PoL2e.com/mc23.14
Lungfishes were important predators in shallow-water habitats in the Devonian, but most lineages died out. The six surviving species live in stagnant swamps and muddy waters in South America, Africa, and Australia (FIGURE 23.43B). Lungfishes have lungs derived from the lunglike sacs of their ancestors as well as gills. When ponds dry up, individuals of most species can burrow deep into the mud and survive for many months in an inactive state while breathing air.
506
Evidence suggests that some early aquatic lobe-limbed vertebrates began to use terrestrial food sources, became more fully adapted to life on land, and eventually evolved to become ancestral tetrapods (“four legs”). How was this transition from an animal that swam in water to one that walked on land accomplished? Fossil evidence suggests that limbs able to prop up a large fish and make the front-to-rear movements necessary for walking evolved while these animals still lived in water. These limbs appear to have functioned in holding the animals upright in shallow water, perhaps even allowing them to hold the head above the water’s surface. These same structures were then co-opted for movement on land, at first probably for foraging on brief trips out of water (FIGURE 23.44). The basic skeletal elements of those limbs can be traced through major changes in limb form and function among the terrestrial vertebrates.
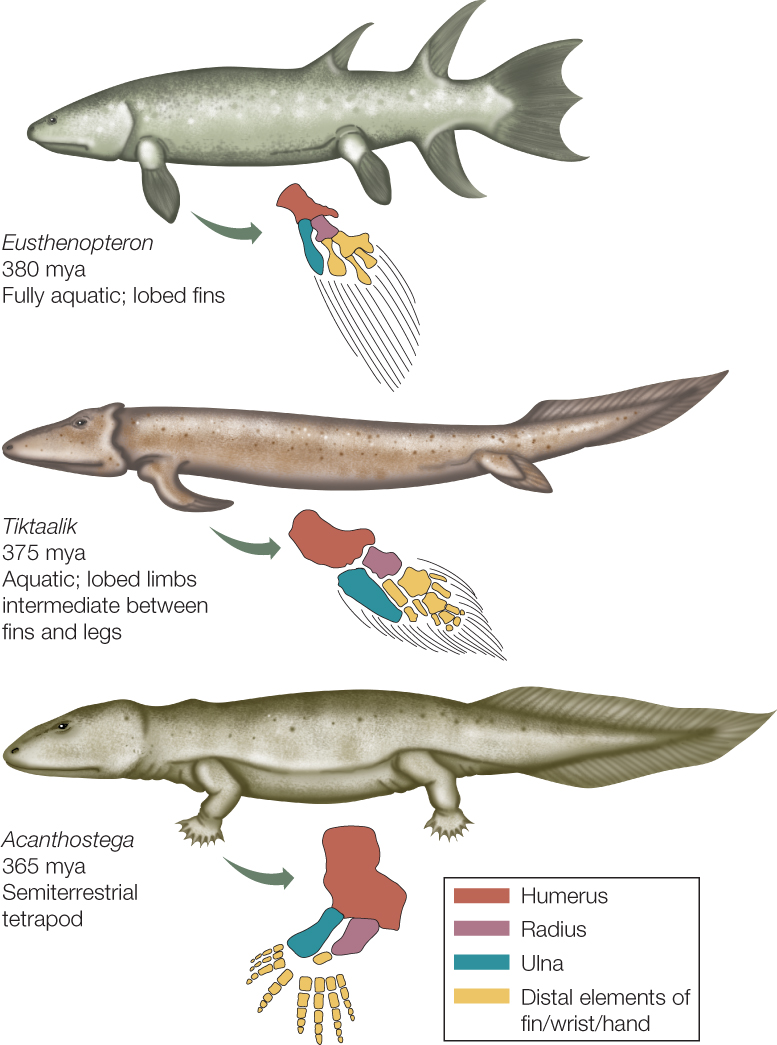
An early split in the tetrapod tree led to two main groups of terrestrial vertebrates: the amphibians, most of which remained tied to moist environments, and the amniotes, many of which adapted to much drier conditions.
Amphibians adapted to life on land
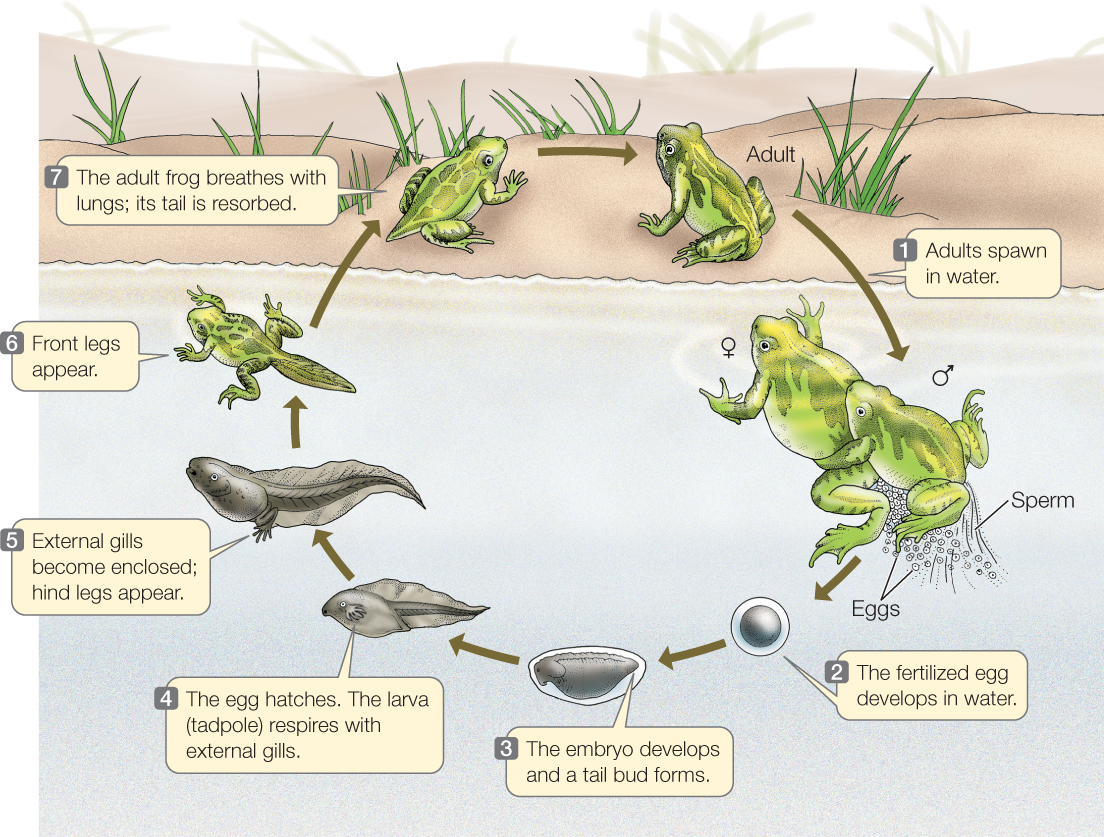
Most modern amphibians are confined to moist environments because they lose water rapidly through the skin when exposed to dry air. In addition, their eggs are enclosed within delicate membranous envelopes that cannot prevent water loss in dry conditions. In some amphibian species, adults live mostly on land but return to fresh water to lay and fertilize their eggs (FIGURE 23.45). The fertilized eggs give rise to larvae that live in water until they undergo metamorphosis to become terrestrial adults. However, many amphibians (especially those in tropical and subtropical areas) have evolved a wide diversity of additional reproductive modes and types of parental care. Internal fertilization, for example, evolved many times among the amphibians. Many species develop directly into adultlike forms from fertilized eggs laid on land or carried by the parents. Other species of amphibians are entirely aquatic, never leaving the water at any stage of their lives, and many of these species retain a larval-like morphology.

Go to ANIMATED TUTORIAL 23.4 Life Cycle of a Frog
PoL2e.com/at23.4
The more than 7,000 known species of amphibians living on Earth today belong to three major groups: the wormlike, limbless, tropical, burrowing or aquatic caecilians (FIGURE 23.46A), the tailless frogs and toads (collectively called anurans; FIGURE 23.46B), and the tailed salamanders (FIGURE 23.46C,D).

Anurans are most diverse in wet tropical and warm temperate regions, although a few are found at very high latitudes. There are far more anurans than any other amphibians, with well over 6,000 described species and more being discovered every year. Some anurans have tough skins and other adaptations that enable them to live for long periods in very dry deserts, whereas others live in moist terrestrial and arboreal environments. Some species are completely aquatic as adults. All anurans have a very short vertebral column, with a strongly modified pelvic region that is adapted for leaping, hopping, or propelling the body through water by kicking the hind legs.
The more than 600 described species of salamanders are most diverse in temperate regions of the Northern Hemisphere, but many species are also found in cool, moist environments in the mountains of Central America, and a few species penetrate into tropical regions. Many salamanders live in rotting logs or moist soil. One major group has lost lungs, and these species exchange gases entirely through the skin and mouth lining—body parts that all amphibians use in addition to their lungs. A completely aquatic lifestyle has evolved several times among the salamanders (see Figure 23.46D). These aquatic species have arisen through a developmental process known as neoteny, or the retention of juvenile traits (such as gills) by delayed somatic development. Most species of salamanders have internal fertilization, which is usually achieved through the transfer of a small jellylike, sperm-embedded capsule called a spermatophore.
507
Many amphibians have complex social behaviors. Most male anurans utter loud, species-specific calls to attract females of their own species (and sometimes to defend breeding territories), and they compete for access to females that arrive at the breeding sites. Many amphibians lay large numbers of eggs, which they abandon once they are deposited and fertilized. Some amphibians lay only a few eggs, which are fertilized and then guarded in a nest or carried on the backs, in the vocal pouches, or even in the stomachs of one of the parents. A few species of frogs, salamanders, and caecilians are viviparous, meaning they give birth to well-developed young that have received nutrition from the female during gestation.
508

Go to MEDIA CLIP 23.15 Answering a Mating Call
PoL2e.com/mc23.15
Amphibians are the focus of much attention today because populations of many species are declining rapidly, especially in mountainous regions of western North America, Central and South America, and northeastern Australia. Worldwide, about one-third of amphibian species are now threatened with extinction or have disappeared completely in the last few decades. Scientists are investigating several hypotheses to account for these population declines, as described in Chapters 1 and 22.
Amniotes colonized dry environments
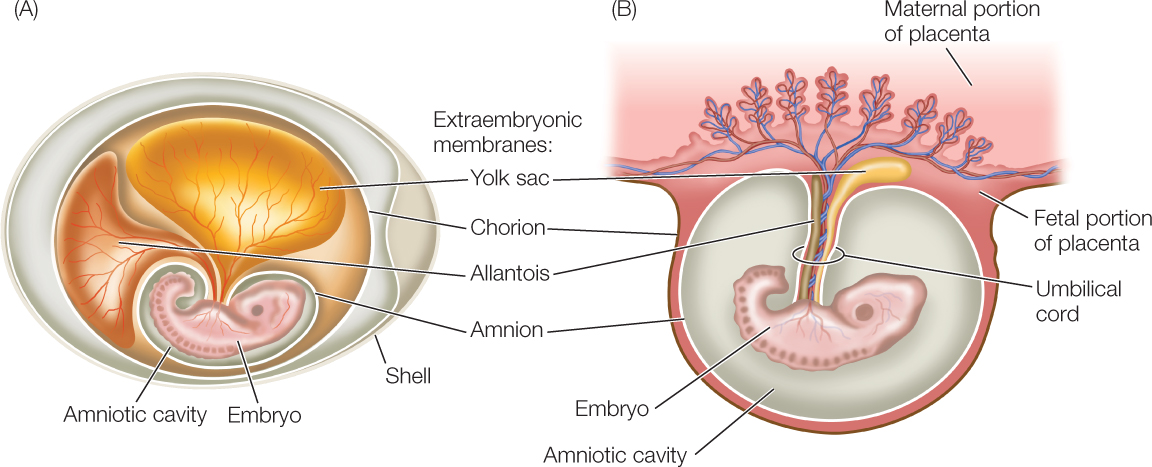
Several key innovations contributed to the ability of amniotes to exploit a wide range of terrestrial habitats. The amniote egg (which gives the group its name) is relatively impermeable to water and allows the embryo to develop in a contained aqueous environment (FIGURE 23.47A). The leathery or brittle, calcium-impregnated shell of the amniote egg retards evaporation of the fluids inside but permits passage of oxygen and carbon dioxide. The egg also stores large quantities of food in the form of yolk, allowing the embryo to attain a relatively advanced state of development before it hatches. Within the shell are extraembryonic membranes that protect the embryo from desiccation and assist its gas exchange and excretion of waste nitrogen.
Go to ACTIVITY 23.3 The Amniote Egg
PoL2e.com/ac23.3
LINK
The roles of the extraembryonic membranes of the amniote egg are discussed in Concept 38.5
In several different groups of amniotes, the amniote egg became modified, allowing the embryo to grow inside (and receive nutrition from) the mother. For instance, the mammalian egg lost its shell whereas the functions of the extraembryonic membranes were retained and expanded (FIGURE 23.47B).
Other innovations evolved in the organs of terrestrial adults. A tough, impermeable skin, covered with scales or modifications of scales such as hair and feathers, greatly reduced water loss. Adaptations of the vertebrate excretory organs, the kidneys, allowed amniotes to excrete concentrated urine, ridding the body of waste nitrogen without losing a large amount of water in the process.
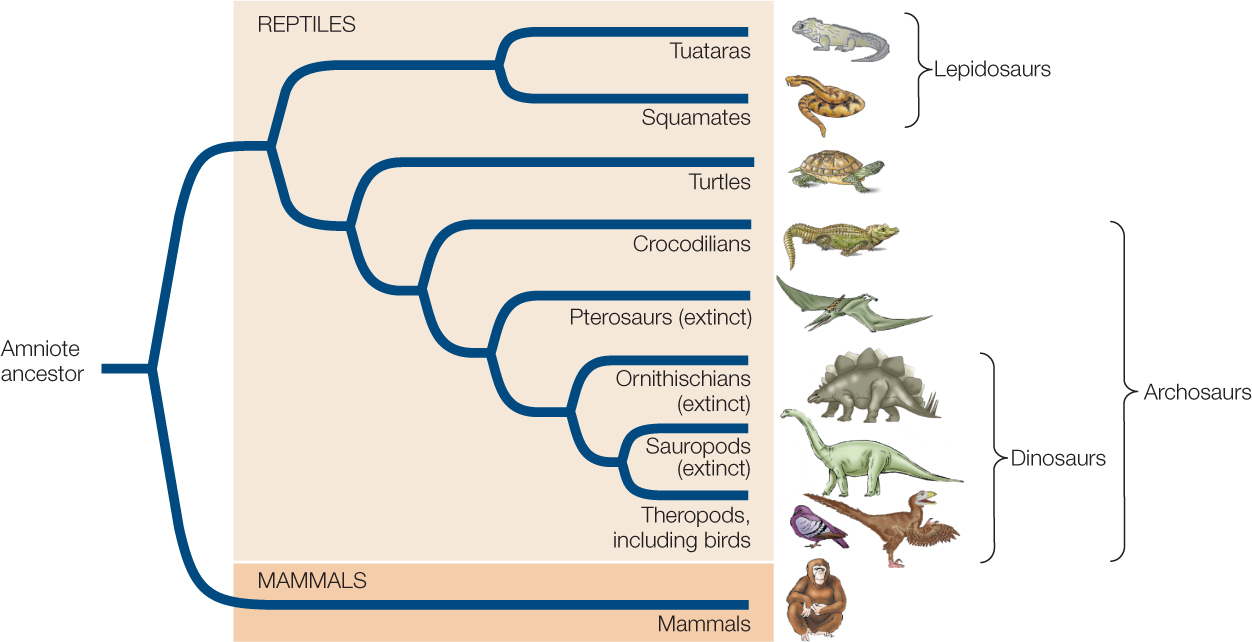
During the Carboniferous, amniotes split into two major groups: the reptiles and the lineage that eventually led to the mammals (FIGURE 23.48).
Reptiles adapted to life in many habitats
The lineage leading to modern reptiles began to diverge from other amniotes more than 300 million years ago. More than 19,000 species of reptiles exist today, more than half of which are birds. Birds are the only living members of the otherwise extinct dinosaurs, the dominant terrestrial predators of the Mesozoic.
The lepidosaurs constitute the second-most species-rich clade of living reptiles. This group is composed of the squamates (lizards, snakes, and amphisbaenians—the last a group of mostly legless, wormlike, burrowing reptiles with greatly reduced eyes) and the tuataras, which superficially resemble lizards but differ from them in tooth attachment and several internal anatomical features. Many species related to the tuataras lived during the Mesozoic era, but today only two species, restricted to a few islands off New Zealand, survive (FIGURE 23.49A).

The skin of a lepidosaur is covered with horny scales that greatly reduce loss of water from the body surface. These scales, however, make the skin unavailable as an organ of gas exchange. Gases are exchanged almost entirely via the lungs, which are proportionally much larger in surface area than those of amphibians. A lepidosaur forces air into and out of its lungs by bellowslike movements of its ribs. The three-chambered lepidosaur heart partially separates oxygenated blood from the lungs from deoxygenated blood returning from the body. With this type of heart, lepidosaurs can generate high blood pressure and can sustain a relatively high metabolism.
509
510
Most lizards are insectivores, but some are herbivores; a few prey on other vertebrates. Many lizards walk on four limbs (FIGURE 23.49B), although limblessness has evolved repeatedly in the group, especially in burrowing and grassland species. The largest lizard, which grows as long as 3 meters and can weigh more than 150 kilograms, is the predaceous Komodo dragon of the East Indies.

Go to MEDIA CLIP 23.16 Komodo Dragons Bring Down Prey
PoL2e.com/mc23.16
One major group of limbless squamates is the snakes (FIGURE 23.49C). All snakes are carnivores, and many can swallow objects much larger than themselves. Several snake groups evolved venom glands and the ability to inject venom rapidly into their prey.
The turtles comprise a reptilian group that has changed relatively little since the early Mesozoic. In these reptiles, dorsal and ventral bony plates form a shell into which the head and limbs can be withdrawn in many species (FIGURE 23.49D). The dorsal shell is a modification of the ribs. It is a mystery how the pectoral girdles evolved to be inside the ribs of turtles, making them unlike any other vertebrates. Most turtles live in aquatic environments, but several groups, such as tortoises and box turtles, are terrestrial. Sea turtles spend their entire lives at sea except when they come ashore to lay eggs. Human exploitation of sea turtles and their eggs has resulted in worldwide declines of these species, all of which are now endangered. A few species of turtles are strict herbivores or carnivores, but most species are omnivores that eat a variety of aquatic and terrestrial plants and animals.
Crocodilians and birds share their ancestry with the dinosaurs
Another reptilian clade, the archosaurs, includes the crocodilians, pterosaurs, dinosaurs, and birds. Only the crocodilians and birds are represented by living species today. Modern crocodilians—crocodiles, caimans, gharials, and alligators—are confined to tropical and warm temperate environments (FIGURE 23.50A). All crocodilians are carnivorous; they eat vertebrates of all kinds, including large mammals. Crocodilians spend much of their time in water, but they lay their eggs in nests they build on land or on floating piles of vegetation. The eggs are warmed by heat generated by decaying organic matter that the female places in the nest. The female provides other forms of parental care as well: typically, she guards the eggs until they hatch, and in some species, she continues to guard and communicate with her offspring after they hatch.

Dinosaurs rose to prominence about 215 million years ago and dominated terrestrial environments for about 150 million years. However, only one group of dinosaurs, the birds, survived the mass extinction at the end of the Cretaceous. During the Mesozoic, most terrestrial animals more than a meter long were dinosaurs. Many were agile and could run rapidly; they had special muscles that enabled the lungs to be filled and emptied while the limbs moved. We can infer the existence of such muscles in dinosaurs from the structure of the vertebral column in fossils. Some of the largest dinosaurs weighed as much as 70,000 kilograms.
Biologists have long accepted the phylogenetic position of birds among the reptiles, although birds clearly have many unique, derived morphological features. In addition to the strong morphological evidence for this placement, fossil and molecular data emerging over the last few decades have provided definitive supporting evidence. Birds are a specialized group of theropods, a clade of predatory dinosaurs that shared such traits as a bipedal stance, hollow bones, a furcula (“wishbone”), elongated metatarsals with three-fingered feet, elongated forelimbs with three fingers, and a pelvis that points backward. Modern birds are homeothermic, meaning they regulate their body temperatures at a relatively constant temperature through physiological control. Although we cannot directly assess this physiological trait in extinct species, many fossil theropods share morphological traits that suggest they may have been homeothermic as well.
511
LINK
Homeothermy and its physiological control are described in Concept 29.3
The living bird species fall into two major groups that diverged about 80–90 million years ago from a flying ancestor. The few modern descendants of one lineage include a group of secondarily flightless and weakly flying birds, some of which are very large. This group, called the palaeognaths, includes the South and Central American tinamous and several large flightless birds of the southern continents—the rheas, emu, kiwis, cassowaries, and the world’s largest bird, the ostrich (FIGURE 23.50B). The second lineage, the neognaths, has left a much larger number of descendants, most of which have retained the ability to fly.
The evolution of feathers allowed birds to fly
During the Mesozoic era, about 175 million years ago, a lineage of theropods gave rise to the birds. Recent fossil discoveries show that the scales of some theropod dinosaurs were modified to form feathers. The feathers of many of these dinosaurs were structurally similar to those of modern birds (FIGURE 23.51).

The evolution of feathers was a major force for diversification. Feathers are lightweight but are strong and structurally complex. The large quills of the flight feathers on the wings arise from the skin of the forelimbs to create the flying surfaces. Other strong feathers sprout like a fan from the shortened tail and serve as stabilizers during flight. The feathers that cover the body, along with an underlying layer of down feathers, provide birds with insulation that helps them survive in virtually all of Earth’s climates.

Go to MEDIA CLIP 23.17 Falcons in Flight
PoL2e.com/mc23.17
The bones of theropod dinosaurs, including birds, are hollow with internal struts that increase their strength. Hollow bones would have made early theropods lighter and more mobile. Later, the hollow bones of theropods facilitated the evolution of flight. The sternum (breastbone) of flying birds forms a large, vertical keel to which the flight muscles are attached.
Flight is metabolically expensive. A flying bird consumes energy at a rate 15–20 times faster than a running lizard of the same weight. Because birds have such high metabolic rates, they generate large amounts of heat. They control the rate of heat loss using their feathers, which may be held close to the body or elevated to alter the amount of insulation they provide. The lungs of birds allow air to flow through unidirectionally rather than by pumping air in and out. This flow-through structure of the lungs increases the efficiency of gas exchange and thereby supports an increased metabolic rate.
LINK
You can read more about the effects of flying on metabolic rate in Concept 29.2. See Concept 31.2 for a description of the breathing system of birds
There are about 10,000 species of living birds, which range in size from the 150-kilogram ostrich to a hummingbird weighing only 2 grams (FIGURE 23.52). The teeth so prominent among other dinosaurs were secondarily lost in the ancestral birds, but birds nonetheless eat almost all types of animal and plant material. Insects and fruits are the most important dietary items for terrestrial species. Birds also eat seeds, nectar and pollen, leaves and buds, carrion, and fish and other vertebrates. By eating the fruits and seeds of plants, birds serve as major agents of seed dispersal.

Mammals radiated as non-avian dinosaurs declined in diversity
Small and medium-sized mammals coexisted with the large dinosaurs throughout most of the Mesozoic era, and most of the major groups that are alive today arose in the Cretaceous. After the non-avian dinosaurs disappeared during the mass extinction at the end of the Cretaceous, mammals increased dramatically in numbers, diversity, and size. Today mammals range in size from tiny shrews and bats weighing only about 2 grams to the blue whale, the largest animal on Earth, which measures up to 33 meters long and can weigh as much as 160,000 kilograms. Mammals have far fewer, but more highly differentiated, teeth than do fishes, amphibians, or reptiles. Differences among mammals in the number, type, and arrangement of teeth reflect their varied diets.
512
Four key features distinguish the mammals:
- Sweat glands, which secrete sweat that evaporates and thereby cools an animal
- Mammary glands, which in females secrete a nutritive fluid (milk) on which newborn individuals feed
- Hair, which provides a protective and insulating covering
- A four-chambered heart that completely separates the oxygenated blood coming from the lungs from the deoxygenated blood returning from the body (this last characteristic evolved independently in the archosaurs, including modern birds and crocodiles)
Mammalian eggs are fertilized within the female’s body, and the embryos undergo a period of development in the female’s body in an organ called the uterus. In the uterus, the embryo is contained in an amniotic sac that is homologous to one of four membranes found in the amniotic egg (see Figure 23.47). The embryo is connected to the wall of the uterus by an organ called a placenta. The placenta allows for nutrient and gas exchange, as well as waste elimination from the developing embryo, via the female’s circulatory system. Most mammals have a covering of hair (fur), which is luxuriant in some species but has been greatly reduced in others, including the cetaceans (whales and dolphins) and humans. Thick layers of insulating fat (blubber) replace hair as a heat-retention mechanism in the cetaceans. Humans learned to use clothing for heat retention when they dispersed from warm tropical areas.
The approximately 5,700 species of living mammals are divided into two primary groups: the prototherians and the therians. Only five species of prototherians are known, and they are found only in Australia and New Guinea. These mammals, the duck-billed platypus and four species of echidnas, differ from other mammals in lacking a placenta, laying eggs, and having sprawling legs (FIGURE 23.53A). Prototherians supply milk for their young, but they have no nipples on their mammary glands. The milk simply oozes out and is lapped off the fur by the offspring.

Most mammals are viviparous
Members of the viviparous therian clade are further divided into the marsupials and the eutherians. Females of most marsupial species have a ventral pouch in which they carry and feed their offspring (FIGURE 23.53B). Gestation (pregnancy) in marsupials is brief; the young are born tiny but with well-developed forelimbs, with which they climb to the pouch. They attach to a nipple but cannot suck. The mother ejects milk into the tiny offspring until they grow large enough to suckle. Once her offspring have left the uterus, a female marsupial may become sexually receptive again. She can then carry fertilized eggs that are capable of initiating development and can replace the offspring in her pouch should something happen to them.
Eutherians include the majority of mammals. Eutherians are sometimes called placental mammals, but this name is inappropriate because some marsupials also have placentas. Eutherians are more developed at birth than are marsupials, and no external pouch houses them after they are born.
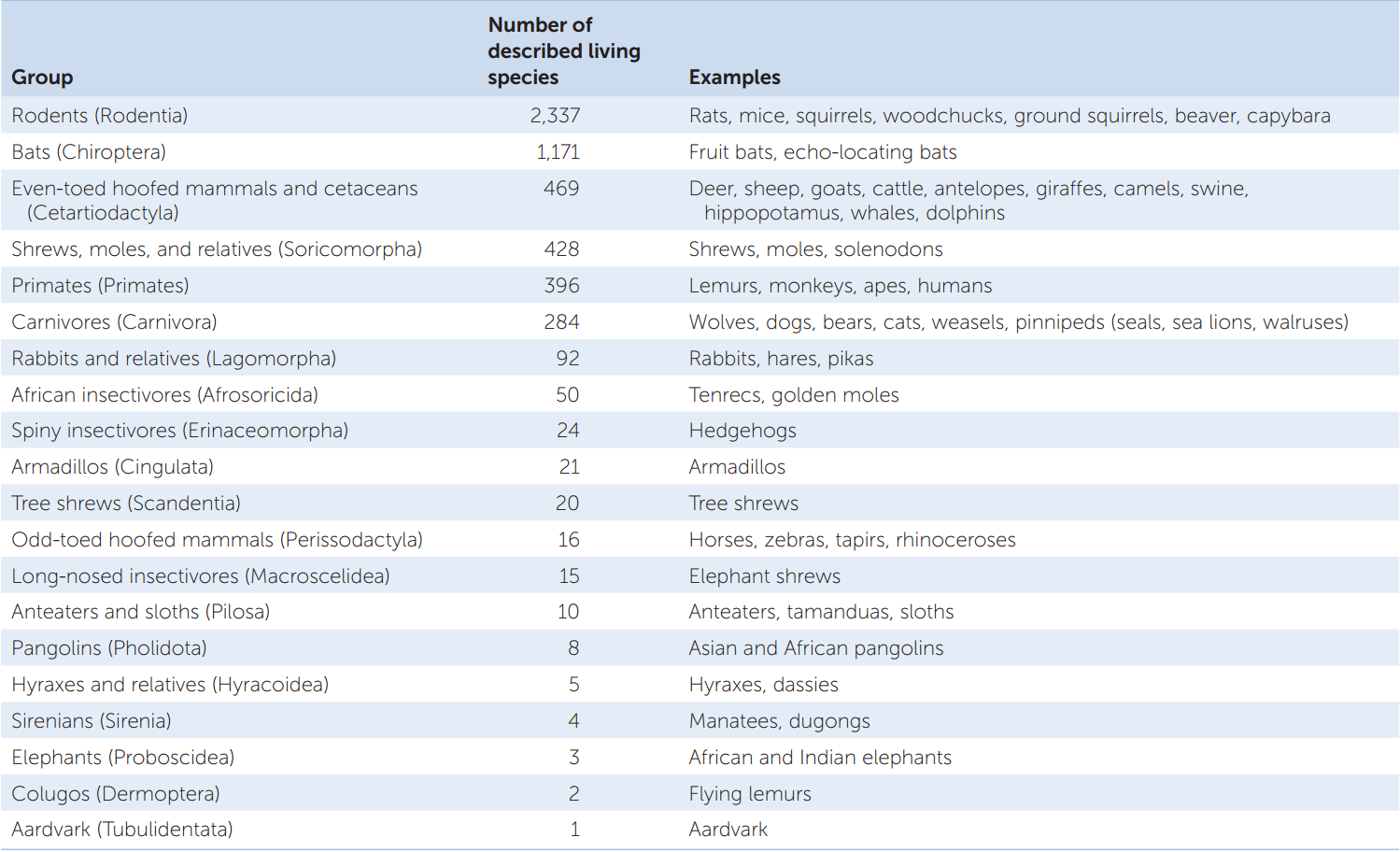
The more than 5,300 living species of morphologically diverse eutherians are divided into 20 major groups (TABLE 23.3). The relationships of the major groups of eutherians to one another have been difficult to determine because most of the major groups diverged in a short period of time during an explosive adaptive radiation. The two most diverse groups are the rodents (FIGURE 23.53C) and bats (FIGURE 23.53D), which together comprise about two-thirds of the species. Rodents are traditionally defined by the unique morphology of their teeth, which are adapted for gnawing through substances such as wood. The bats probably owe much of their success to the evolution of flight, which allows them to exploit a variety of food sources and colonize remote locations with relative ease.
513

Go to MEDIA CLIP 23.18 Bats Feeding in Flight
PoL2e.com/mc23.18
Grazing and browsing by members of several eutherian groups helped transform the terrestrial landscape. Herds of grazing herbivores feed on open grasslands, whereas browsers feed on shrubs and trees. The effects of herbivores on plant life favored the evolution of the spines, tough leaves, and difficult-to-eat growth forms found in many plants. In turn, adaptations to the teeth and digestive systems of many herbivore lineages allowed these species to consume many plants despite such defenses—a striking example of coevolution. A large animal can survive on food of lower quality than a small animal can, and large size evolved in several groups of grazing and browsing mammals (FIGURE 23.53E). The evolution of large herbivores, in turn, favored the evolution of large carnivores able to attack and overpower them.
514
Several lineages of terrestrial eutherians subsequently returned to the aquatic environments their ancestors had left behind (FIGURE 23.53F). The completely aquatic cetaceans—whales and dolphins—evolved from artiodactyl ancestors (whales are closely related to the hippopotamuses). The seals, sea lions, and walruses also returned to the marine environment, and their limbs became modified into flippers. Weasel-like otters retain their limbs but have also returned to aquatic environments, colonizing both fresh and salt water. The manatees and dugongs colonized estuaries and shallow seas.
CHECKpoint CONCEPT 23.6
- Many amphibians have a biphasic life cycle that includes an aquatic larva and a terrestrial adult. What are some exceptions to this common pattern?
- In the not-too-distant past, the idea that birds were reptiles met with skepticism. Explain how fossils, morphology, and molecular evidence now support the position of birds among the reptiles.
- Contrast the reproductive modes of prototherians, marsupials, and eutherians.
The biology of one eutherian group—the primates—has been the subject of extensive research. The behavior, ecology, physiology, and molecular biology of the primates are of special interest to us because this lineage includes humans.