Concept 25.2: Soil Organisms Contribute to Plant Nutrition
One gram of soil contains from 6,000 to 50,000 bacterial species and up to 200 meters of fungal hyphae, although both are largely invisible to the naked eye. Many of these soil organisms consume living and dead plant materials. Plants have an array of defenses they can use against potentially harmful soil organisms (which we will describe in Chapter 28). In many cases, however, plants actively encourage fungi and bacteria to infect their roots. Here we describe the resulting “intercellular trading posts,” where products are exchanged to the mutual benefit of plants and a few very special soil microbes. In these interactions—called symbioses (singular symbiosis)—the plants usually “trade” the products of photosynthesis, which the microbes use as an energy source, for other essential elements, such as phosphorus and nitrogen, which the microbes can supply.
542
LINK
These plant–microbe interactions are examples of mutualism, one of five major types of interspecies interactions discussed in Concept 43.1
We will focus on two important plant–microbe interactions:
- Arbuscular mycorrhizae, a plant–fungus symbiosis (see Figure 22.10)
- Symbiotic nitrogen fixation, an association of nitrogen-fixing bacteria (see Concept 19.3) with plant roots
We’ll begin by describing how these associations form. Then we’ll see how they benefit the plants and their microbial partners.
Plants send signals for colonization
In the case of both arbuscular mycorrhizae and symbiotic nitrogen fixation, the roots of the potential host plant secrete signaling molecules that attract the soil organisms. These signals contribute to the specificity of the interactions, because only one or a few microbe species will respond to a particular signal molecule.
Formation of Mycorrhizal Associations
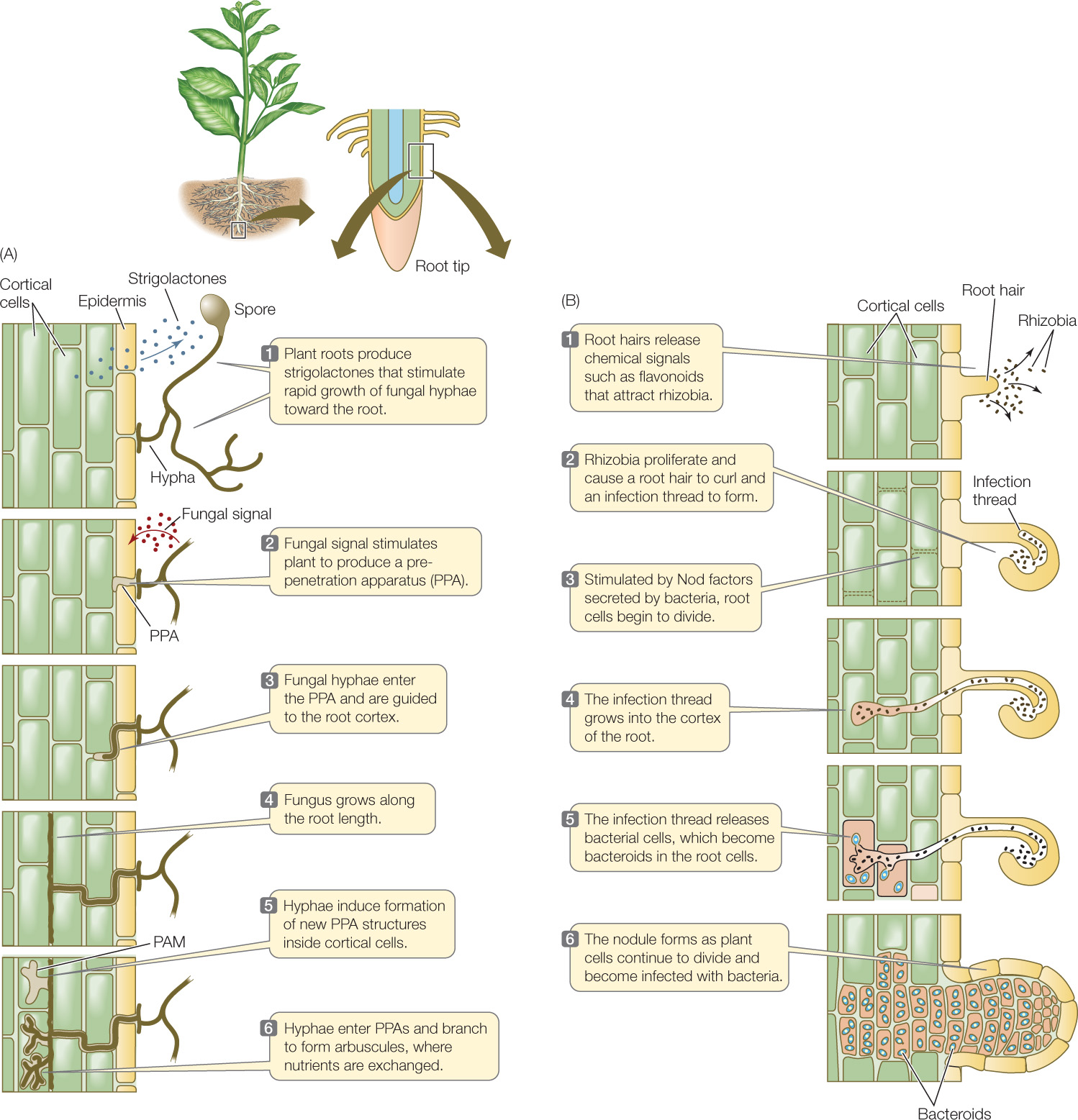
In the case of arbuscular mycorrhizae, plant roots produce chemical signal molecules called strigolactones that stimulate rapid growth of fungal hyphae toward the root (FIGURE 25.5A). In response, the fungi signal the plant to form a prepenetration apparatus (PPA), which guides the growth of the fungal hyphae into the root cortex. The sites of nutrient exchange between fungus and plant are the arbuscules, which form inside root cortical cells. Despite the intimacy of this association, the plant and fungal cytoplasms never mix—they are separated by two membranes, the fungal cell membrane and the periarbuscular membrane (PAM), which is continuous with the plant cell membrane.
Formation of Nitrogen-Fixing Nodules
A group of plants called legumes (family Fabaceae) can form symbioses with several species of soil bacteria collectively known as rhizobia, which live in nodules that form on the legumes’ roots. The roots of legumes release flavonoids and other chemical signals that attract the rhizobia (FIGURE 25.5B). The flavonoids also stimulate the bacteria to secrete Nod (nodulation) factors, which cause cells in the root cortex to divide and form the plant tissue that constitutes the root nodule. Bacteria enter the root via an infection thread, analogous to the PPA in mycorrhizal associations, and eventually reach cells inside the root nodule. There the bacteria are released into the cytoplasm of the nodule cells, enclosed in plant membrane–derived vesicles that are functionally equivalent to the PAM. The bacteria differentiate into bacteroids—the form of the bacteria that can fix nitrogen.
A Common Mechanism
There is increasing evidence that nodule formation depends on some of the same genes and mechanisms that allow mycorrhizal associations to develop. For example, both processes involve folding in of the cell membrane to allow entry of the fungal hyphae or rhizobia. The similarities in the structures formed (see Figure 25.5) are striking, considering that the symbioses involve members of two different groups of organisms (fungi and bacteria).
LINK
The fact that similar genes and developmental processes are used in two different types of symbiosis provides a striking demonstration of the principles of evolutionary developmental biology; see Concept 14.4
Mycorrhizae expand the root system
In most vascular plants, the roots alone cannot support an optimal growth rate—they simply cannot reach all the nutrients available in the soil. Mycorrhizae, which occur in more than 90 percent of terrestrial plants, expand the root surface area 10-fold to 1,000-fold (see Figure 22.10A). Fungal hyphae are also much finer than root hairs; they can get into pores in the soil that are inaccessible to roots. Mycorrhizae thus probe a vast expanse of soil for nutrients and deliver them into root cortical cells.
APPLY THE CONCEPT: Soil organisms contribute to plant nutrition
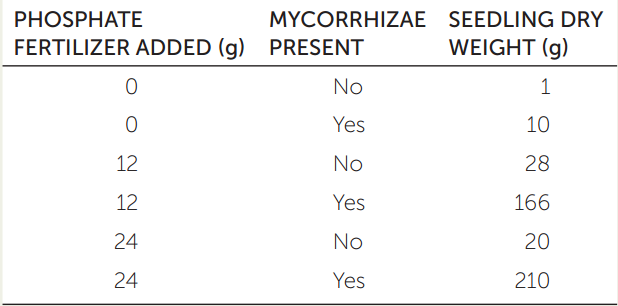
Researchers grew lemon seedlings in soils that contained phosphate fertilizer, arbuscular mycorrhizal fungi, or both. The table shows the mean dry weight of the seedlings after 6 months.
- What do these results tell us about the role of phosphate fertilizer in the growth of lemon plants?
- Why didn’t the mean weight of the seedlings increase when the amount of phosphate fertilizer, in the absence of mycorrhizae, was doubled?
543
The primary nutrient that the plant obtains from a mycorrhizal interaction is phosphorus. In exchange, the fungus obtains an energy source: the products of photosynthesis. In fact, up to 20 percent of the photosynthate (carbohydrates produced by photosynthesis) of terrestrial plants is directed to and consumed by arbuscular mycorrhizal fungi.
Rhizobia capture nitrogen from the air and make it available to plant cells
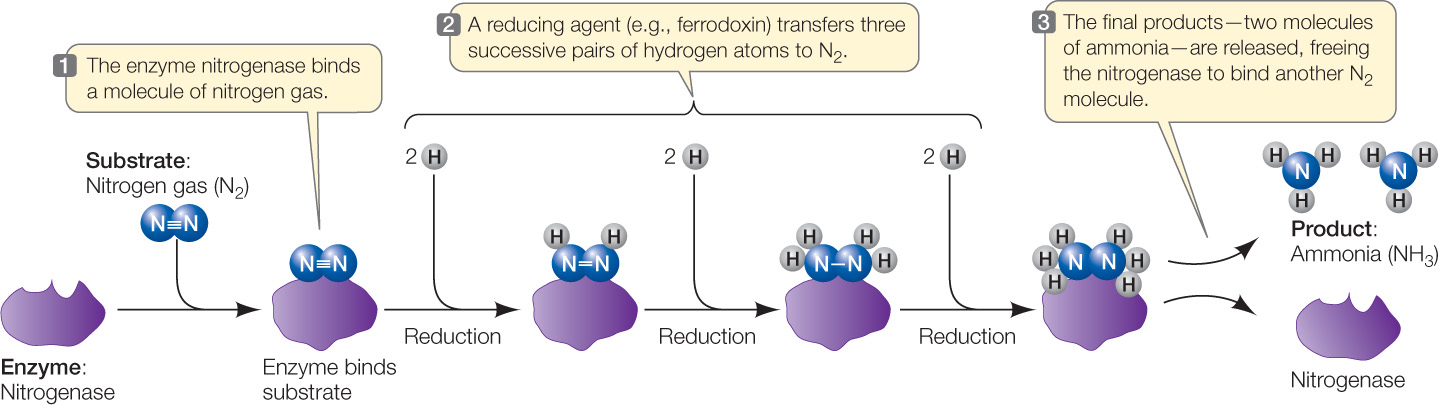
Although nitrogen is essential for the survival of plants, they cannot use atmospheric N2 because they lack the biochemical machinery to break the very stable triple bond that links the two N atoms in N2. Some bacteria, however, have an enzyme called nitrogenase that enables them to convert N2 into a more reactive and biologically useful form, ammonia (NH3), by a process called nitrogen fixation:
N2 + 6 H → 2 NH3
544
This process is by far the most important route by which nitrogen enters biological systems. Two types of organisms can fix nitrogen:
- Free-living (nonsymbiotic) organisms in soil and water (e.g., Azotobacter bacteria and Nostoc cyanobacteria)
- Symbiotic organisms living in other organisms (e.g., rhizobia in the roots of legumes)
Biological nitrogen fixation is the reduction of nitrogen gas, and it requires a great deal of energy. It proceeds by the stepwise addition of three pairs of hydrogen atoms to N2 (FIGURE 25.6).
LINK
You can review reduction–oxidation reactions in Concept 6.1
Nitrogenase is strongly inhibited by oxygen. Many free-living nitrogen-fixing bacteria are anaerobes that live in environments that naturally contain little or no O2. The environment in the plant root tissues is typically aerobic, but the nodules provide the rhizobia with the conditions necessary for nitrogen fixation (see Figure 25.5B). Within a nodule, O2 is maintained at a low level that is sufficient to support aerobic respiration by the bacteria (which is necessary to supply energy for the fixation reaction) but not so high as to inactivate nitrogenase. The O2 level is regulated by a plant-produced protein called leghemoglobin, which is an O2 carrier. Leghemoglobin is a close relative of hemoglobin, the red, oxygen-carrying pigment of animals, and is thus an evolutionarily ancient molecule.
As with mycorrhizae, maintaining nitrogen-fixing bacteria is costly to the plant, consuming as much as 20 percent of all of the energy stored in photosynthate. Relatively few plant species are legumes, but those species are important to humans because they can be grown in nitrogen-poor soil. They include peas, soybeans, clover, alfalfa, and some tropical shrubs and trees. Most other plants depend on nitrogen fixed by free-living bacteria in the soil or released by the breakdown of proteins present in dead organic matter.
Some plants obtain nutrients directly from other organisms
Although the majority of plants obtain most of their mineral nutrients from the soil solution (with the help of fungi), some, such as carnivorous and parasitic plants, use other sources.
Carnivorous Plants
There are about 500 carnivorous plant species that obtain some of their nutrients by digesting arthropods. These plants typically grow in boggy soils where little nitrogen or phosphorus is available. Digestion (hydrolysis) of arthropod prey helps provide those missing nutrients. A well-known example of a carnivorous plant is the Venus flytrap (genus Dionaea; FIGURE 25.7A), which has a modified leaf with two halves that fold together. When an insect touches trigger hairs on the leaf, its two halves quickly come together, and their spiny margins interlock and trap the insect before it can escape. The leaf then secretes enzymes that digest the prey.

Parasitic Plants
Approximately 1 percent of flowering plant species derive some or all of their water, nutrients, and even photosynthate from other plants. Hemiparasites can photosynthesize but derive water and mineral nutrients from the living bodies of other plants. Mistletoes, for example, carry out some photosynthesis, but they parasitize other plants for water and mineral nutrients and may derive some photosynthetic products from them as well. Dwarf mistletoe (Arceuthobium americanum) is a serious parasite in forests of the western United States, destroying more than 3 billion board feet of lumber per year. Holoparasites are completely parasitic and do not perform photosynthesis. Witchweed (Striga) is a serious parasite of crops, causing more than $1 billion a year in corn losses in Africa (FIGURE 25.7B).
A Common Association
Interestingly, strigolactone, the molecule made by plant roots that attracts arbuscular mycorrhizal fungi, is closely related to the molecule that Striga is attracted to as it seeks potential hosts. The mycorrhizal association is ancient (more than 400 million years old) and predates the evolution of parasitic plants. Scientists hypothesize that the ancestors of modern Striga evolved to take advantage of a compound that was already produced by plants to attract beneficial soil microbes. This is an example of “opportunistic evolution,” the repurposing of preexisting processes rather than the development of new processes.
545
CHECKpoint CONCEPT 25.2
- Compare the events that occur when arbuscular mycorrhizal fungi and rhizobia infect a plant root. What is the nutritional advantage to the plant in each case?
- How might you turn wheat, a plant that does not have symbiotic relationships with nitrogen-fixing bacteria, into a nitrogen-fixing plant? What biochemical challenges would you face?
- How do the nutritional needs of holoparasitic plants differ from those of carnivorous plants?
- What characteristics are shared among nonparasitic plant–parasitic plant, plant–mycorrhizal fungus, and plant–rhizobia associations?
Now that we have learned about plant nutrients in the soil, let’s consider how soil water, with its dissolved nutrients, enters the plant root, and how it moves from the root to the rest of the plant body.