Concept 28.3: Plants Adapt to Environmental Stresses
In an ever-changing environment, plants face several potential stressors: drought, submersion, heat, cold, and high concentrations of salt and heavy metals in the soil. Here we will focus on the overall mechanisms by which plants deal with extremes of climate and soil conditions. You will see that in these cases, as with defenses against pathogens and herbivores, there are constitutive defenses as well as induced responses.

Go to MEDIA CLIP 28.1 Leaves for Every Environment
PoL2e.com/mc28.1
Some plants have special adaptations to live in very dry conditions
Many plants, especially those living in deserts, must cope with extremely limited water supplies. A variety of anatomical and life-cycle adaptations allow plants to survive under these conditions. Many of these adaptations are ways to avoid or reduce the inevitable water loss through transpiration that occurs during active photosynthesis (see Concept 25.3). Other adaptations help plants tolerate the high levels of light and heat that are often found in deserts.
Drought Avoiders
Some desert plants have no special structural adaptations for water conservation. Instead, these desert annuals, called drought avoiders, simply evade periods of drought. Drought avoiders carry out their entire life cycle—from seed to seed—during a brief period in which rainfall has made the surrounding desert soil sufficiently moist for growth and reproduction (FIGURE 28.6). A different drought avoidance strategy is seen in deciduous perennial plants, particularly in Africa and South America, that shed their leaves in response to drought as a way to conserve water. These plants remain dormant until conditions are again favorable for growth.

598
Leaf Structures
Most desert plants are not drought avoiders, but rather grow in their dry environment year-round. Plants adapted to dry environments are called xerophytes (Greek xeros, “dry”). Three structural adaptations are found in the leaves of many xerophytes:
- Specialized leaf anatomy that reduces water loss
- A thick cuticle and a profusion of trichomes over the leaf epidermis, which retard water loss
- Trichomes that diffract and diffuse sunlight, thereby decreasing the intensity of light impinging on the leaves and the risk of damage to the photosynthetic apparatus by excess light
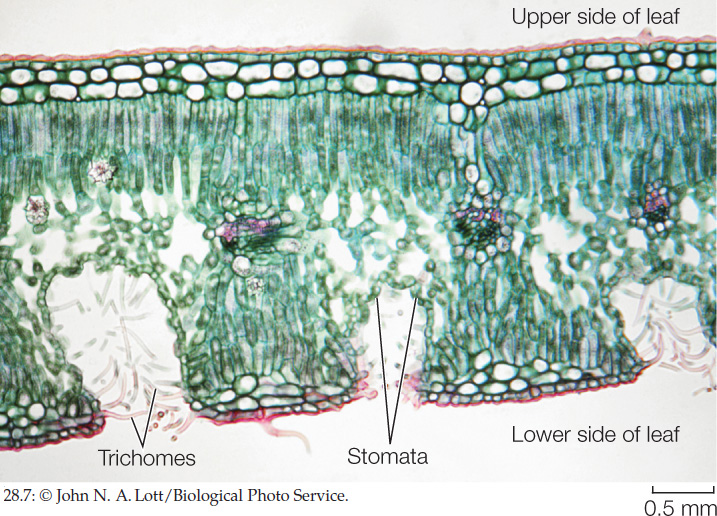
In some xerophytes, the stomata are strategically located in sunken cavities below the leaf surface (known as stomatal crypts), where they are sheltered from the drying effects of air currents (FIGURE 28.7). Trichomes surrounding the stomata slow air currents further. Cacti and similar plants have spines rather than typical leaves, and photosynthesis is confined to the fleshy stems (see Figure 24.13D). The spines may help the plants cope with desert conditions by reflecting solar radiation or by dissipating heat. The spines also deter herbivores.
Water-Storing Structures
Succulence—the possession of fleshy, water-storing leaves or stems—is another adaptation to dry environments (FIGURE 28.8). This adaptation allows plants to take up large amounts of water when it is available (such as after a brief thunderstorm) and then draw on the stored water during subsequent dry periods. Other adaptations of succulents include a reduced number of stomata and a variant form of photosynthesis, both of which reduce water loss.

Root Systems That Maximize Water Uptake
Roots may also be adapted to dry environments. Cacti have shallow but extensive fibrous root systems that effectively intercept water at the soil surface following even light rains. The tamarugo tree (FIGURE 28.9) obtains water through taproots that grow to great depths, reaching water supplies far underground, as well as from condensation on its leaves. The Atacama Desert in northern Chile often goes several years without measurable rainfall, but the landscape there has many surprisingly large shrubs.

Solute Accumulation
Xerophytes and other plants that must cope with inadequate water supplies may accumulate high concentrations of the amino acid proline or of secondary metabolites in their vacuoles. This solute accumulation lowers the water potential in the plant’s cells below that in the soil, which allows the plant to take up water via osmosis. Plants living in saline environments share this and several other adaptations with xerophytes, as we will see shortly.
LINK
Review the principles of water potential and water movement in Concept 25.3
599
Some plants grow in saturated soils
For some plants, the environmental challenge is the opposite of that faced by xerophytes: too much water. They live in environments so wet that the diffusion of oxygen to their roots is severely limited. These plants have shallow root systems that grow slowly; oxygen levels are likely to be highest near the surface of the soil, and slow growth decreases the roots’ need for oxygen.
The root systems of some plants adapted to swampy environments, such as coastal mangrove habitats, have pneumatophores, which are extensions that grow out of the water and up into the air (FIGURE 28.10A). Pneumatophores contain lenticels (openings) that allow oxygen to diffuse through them, aerating the submerged parts of the root system.
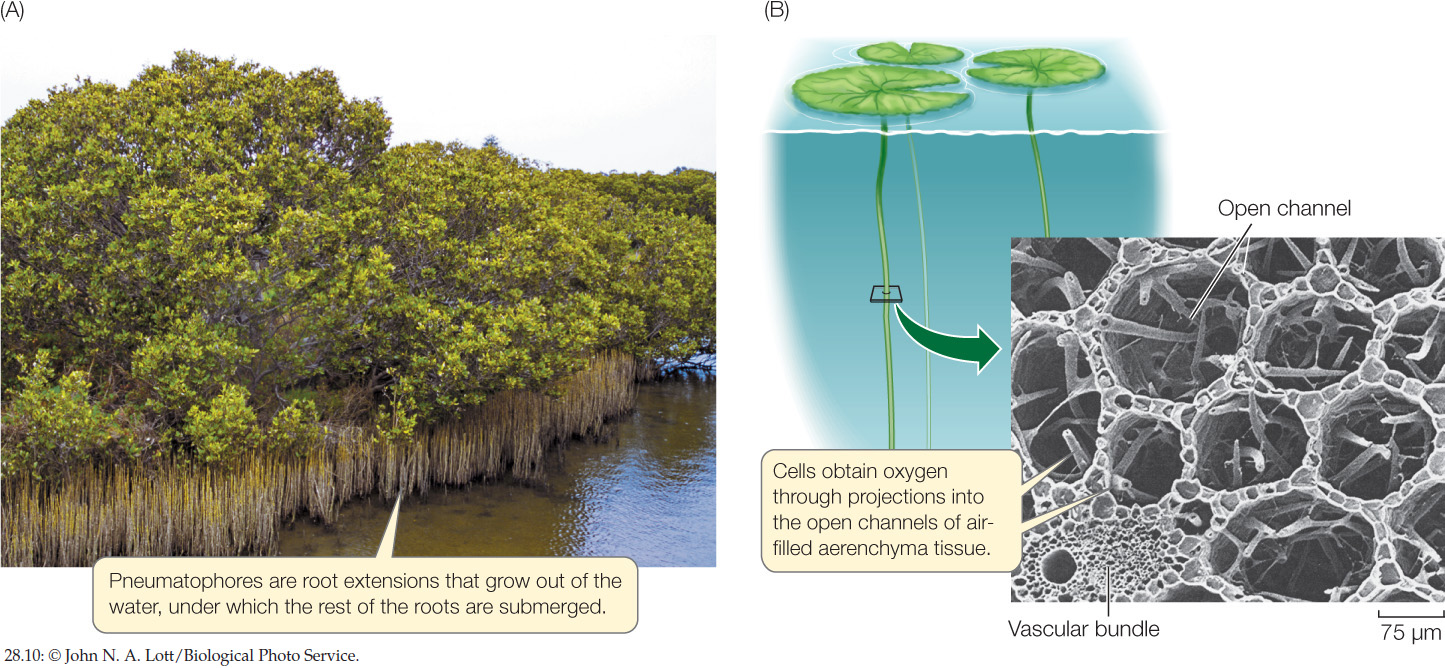
Many submerged or partly submerged aquatic plants have large air spaces in the leaf and stem parenchyma and in the petioles. Tissue containing such air spaces is called aerenchyma (FIGURE 28.10B). Aerenchyma stores oxygen produced by photosynthesis and permits its ready diffusion to parts of the plant where it is needed for cellular respiration. Aerenchyma also imparts buoyancy. Furthermore, because aerenchyma contains far fewer cells than most other plant tissues, metabolism in aerenchyma proceeds at a lower rate, so the need for oxygen is much reduced.
Plants can respond to drought stress
The adaptations of xerophytes for coping with dry environments are generally constitutive—they are always present—and under normal conditions they prevent the plants from experiencing drought stress. When conditions become so dry that even xerophytes are stressed, the plants turn to inducible responses. The same responses are found in many other plants, including those that are not adapted to grow in dry climates.
When the weather is abnormally dry, the water content of the soil is reduced, and less water is available to plants. If a plant cannot take up water from the soil, it cannot grow. Indeed, inadequate water supply is the single most important factor that limits production of our most important food crops. Extreme water deficits in plant cells have two additional major biochemical effects: a reduction in membrane integrity as the polar–nonpolar forces that orient the lipid bilayer are reduced, and changes in the three-dimensional structures of proteins. A plant can suffer irreversible damage when the structure of its cells is compromised in these ways.
When plants sense a water deficit in their roots, a signaling pathway is set in motion that initiates several measures to conserve water and maintain cellular integrity. This pathway begins with the production of the hormone abscisic acid (see Concept 26.3) in the roots. This hormone travels from the roots to the shoot, where it causes stomatal closure and initiates gene transcription that leads to other physiological events that conserve water and cellular integrity (FIGURE 28.11).
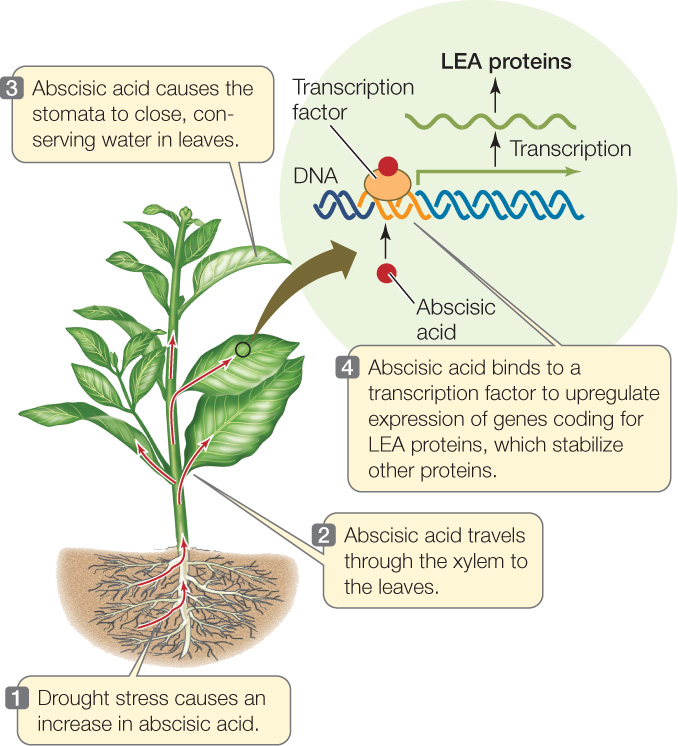
Many plant genes whose expression is altered by drought stress have been identified, largely through research using DNA microarrays, proteomics, and other molecular approaches (see Chapters 12 and 13). One group of proteins in which production is upregulated during drought stress is the late embryogenesis abundant (LEA; pronounced “lee-yuh”) proteins. These hydrophobic proteins also accumulate in maturing seeds as they dry out (hence their name). The LEA proteins bind to membrane proteins and other cellular proteins to stabilize them, preventing them from clumping together as seeds dry out.
600
Plants can cope with temperature extremes
Temperatures that are too high or too low can stress plants and even kill them. Plant species differ in their sensitivity to heat and cold, but all plants have their limits. Any temperature extreme can damage cellular membranes:
- High temperatures destabilize membranes and denature many proteins, especially some of the enzymes of photosynthesis.
- Low temperatures cause membranes to lose their fluidity and alter their permeabilities to solutes.
- Freezing temperatures may cause ice crystals to form, damaging membranes.
Plants have both constitutive defenses and inducible responses for coping with temperature extremes.
Anatomical Adaptations
Many plants living in hot environments have constitutive defenses similar to those of xerophytes. These adaptations include hairs and spines that dissipate heat and leaf forms that intercept less direct sunlight.
Heat Shock Response
The plant inducible response to heat stress is similar to the response to drought stress in that new proteins are made, often under the direction of an abscisic acid–mediated signaling pathway. Within minutes of experimental exposure to raised temperatures (typically a 5°C–10°C increase), plants synthesize several kinds of heat shock proteins. Among these proteins are chaperonins, which help other proteins maintain their structures and avoid denaturation. Threshold temperatures for the production of heat shock proteins vary, but 39°C is sufficient to induce them in most plants.
Cold-Hardening
Low temperatures above the freezing point can cause chilling injury in many plants, including crops such as rice, corn, and cotton as well as tropical plants such as bananas. Many plant species can adjust to cooler temperatures through a process called cold-hardening, which requires repeated exposure to cool temperatures over many days. A key change during the hardening process is an increase in the proportion of unsaturated fatty acids in cell membranes, which allows them to retain their fluidity and function normally at cooler temperatures (see Figure 2.12). Plants have a greater ability to modify the degree of saturation of their membrane lipids than animals do. In addition, low temperatures induce the formation of proteins similar to the LEA proteins related to drought stress and heat shock, which protect against chilling injury (see Figure 28.11).
If ice crystals form within plant cells, they can kill the cells by puncturing organelles and cell membranes. Furthermore, the growth of ice crystals outside the cells can draw water from the cells and dehydrate them. Freeze-tolerant plants have a variety of adaptations to cope with these problems, including the production of antifreeze proteins that slow the growth of ice crystals.
Some plants can tolerate soils with high salt concentrations
Salty, or saline, environments (high in Na+, K+, Ca2+, and Cl−) are found in nature in diverse locales, from hot, dry deserts to moist, cool coastal marshes. Much of the land in Australia is naturally saline. In addition, agricultural land can become increasingly saline as a result of irrigation and the application of chemical fertilizers. This salinization, which can eventually make land unsuitable for farming, is an increasing problem worldwide (FIGURE 28.12).

601
APPLY THE CONCEPT: Plants adapt to environmental stresses
The Arabidopsis gene cor15a is under the control of a promoter that responds to environmental conditions. Arabidopsis plants were genetically modified with a copy of the cor15a promoter fused to a reporter gene that is expressed as an enzyme that is readily detected in leaf extracts (see Concept 13.2 for information about the use of reporter genes to study gene expression). The genetically modified plants were transferred to environments kept at two different temperatures (19°C and 2°C), and the reporter enzyme activity in their leaves was measured at different times after the transfer. The results are shown in the table.a
- Plot these data with hours after transfer on the x axis and enzyme activity on the y axis. What can you conclude about the role of cor15a?
- In a separate experiment, plants were kept at 19°C for 36 hours in the presence of abscisic acid, and the enzyme activity was 8.99. What can you conclude from this result?
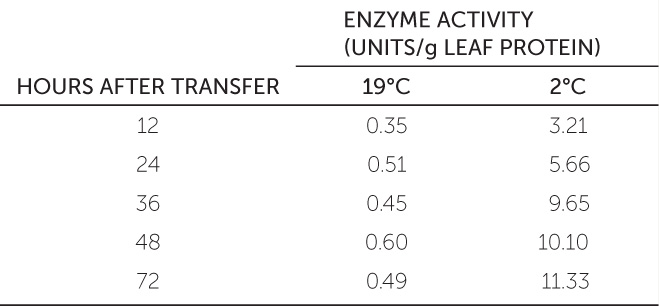
- How would you investigate the possible role of cor15a in the plant’s response to drought? Why would you expect that there might be such a role?
a S. S. Baker et al. 1994. Plant Molecular Biology 24: 701–713.
Because of its high salt concentration, a saline environment has a very negative water potential. To obtain water from such an environment, a plant must have an even more negative water potential (see Concept 25.3); otherwise water will diffuse out of its cells, and the plant will wilt and die. Plants in saline environments are also challenged by the potential toxicity of sodium ions, which inhibit enzymes and protein synthesis.
Plants that are adapted for survival in saline soils are called halophytes. Most halophytes take up Na+, and many take up Cl−, into their roots and transport those ions to their leaves, where they accumulate in the central vacuoles of leaf cells, away from more sensitive parts of the cells. The accumulated salts in the tissues of halophytes make their water potential more negative than the soil solution and allow them to take up water from their saline environment.
Some halophytes have salt glands in their leaves. These glands excrete salt, which collects on the leaf surface until it is removed by rain or wind (FIGURE 28.13). This adaptation, which reduces the danger of poisoning by accumulated salt, is found in some desert plants and in some plants growing in mangrove habitats. The negative water potential in the salt-laden leaves also promotes water flow from the roots up through the xylem by transpiration–cohesion–tension (see Concept 25.3).

Some plants can tolerate heavy metals
High concentrations of heavy metal ions are toxic to most plants. Some soils are naturally rich in heavy metals as a result of normal geological processes or acid rain. The mining of metallic ores also leaves localized areas with high concentrations of heavy metals and low concentrations of nutrients. Such sites are hostile to most plants, and seeds falling on them generally do not produce adult plants.
Some plants can survive in these environments, however, by accumulating concentrations of heavy metals that would kill most plants. More than 200 plant species have been identified as hyperaccumulators that store large quantities of metals such as arsenic (As), cadmium (Cd), nickel (Ni), aluminum (Al), and zinc (Zn).
Perhaps the best-studied hyperaccumulator is alpine pennycress (Thlaspi caerulescens). Before the advent of chemical analysis, miners used the presence of this plant as an indicator of mineral-rich deposits. A Thlaspi plant may accumulate as much as 30 g/kg dry weight Zn (most plants contain 0.1 g/kg dry weight) and 1.5 g/kg dry weight Cd (most plants contain 0.001 g/kg dry weight). Studies of Thlaspi and other hyperaccumulators have revealed the presence of several common adaptations:
- Increased ion transport into the roots
- Increased rates of translocation of ions to the leaves
- Accumulation of ions in vacuoles in the shoot
- Resistance to the ions’ toxicity
Knowledge of these hyperaccumulation mechanisms and the genes underlying them has led to the emergence of phytoremediation, a form of bioremediation (see Chapter 13) that uses plants to clean up environmental pollution in soils. Some phytoremediation projects use natural hyperaccumulators, whereas others use genes from hyperaccumulators to create transgenic plants that grow more rapidly in and are better adapted to a particular polluted environment. In either case, the plants are grown in the contaminated soil, where they act as natural “vacuum cleaners” by taking up the contaminants (FIGURE 28.14). The plants are then harvested and safely disposed of to remove the contaminants. Perhaps the most dramatic use of phytoremediation occurred after an accident at the nuclear power plant at Chernobyl, Ukraine (then part of the Soviet Union), in 1986, when sunflower plants were used to remove uranium from the nearby soil. Phytoremediation is now widely used to clean up land after strip mining.

CHECKpoint CONCEPT 28.3
- Compare the constitutive and induced mechanisms that allow plants to cope with drought conditions.
- Describe a mechanism shared by plants that tolerate high concentrations of salts and those that tolerate heavy metals.
- If an inhibitor of abscisic acid synthesis were sprayed on a plant and the plant was exposed to drought conditions, what would happen?
Question 28.2
How can knowledge of plant and fungal biology be used to prevent the spread of wheat rust?
ANSWER Today, Puccinia graminis strain Ug99 continues its spread and threatens wheat crops worldwide. In the Green Revolution of the 1960s to 1980s, wheat plants were bred to have the gene complexes Sr24 and Sr31, R genes that conferred resistance to existing strains of wheat rust fungus (Concept 28.1). The first strain of Ug99 found in Uganda had Avr genes that form elicitor proteins that do not bind to Sr31 and therefore overcome part of the plant’s resistance. Then, in 2006 a new genetic variant of Ug99 was found in Kenya with additional Avr genes that allow the pathogen to overcome the Sr24 gene resistance as well (FIGURE 28.15).
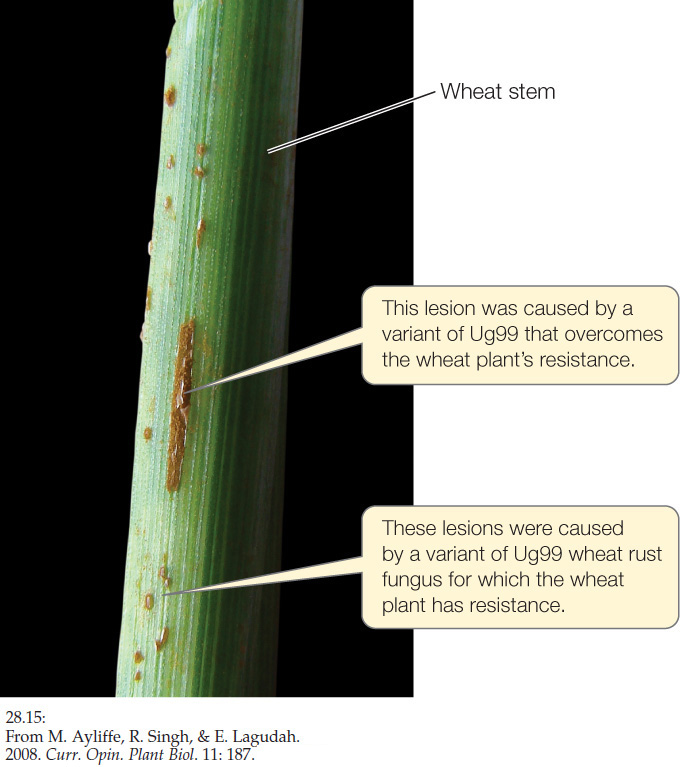
Because 90 percent of the wheat grown in the world has no resistance to this new strain of Ug99, an intensive search is under way for additional wheat resistance genes. In the regions where Ug99 infections are now present, some wheat plants grow that are resistant. Samples of both wheat plants and fungus are sent to laboratories, where, under high security and sterility, they are tested and examined for Avr and R genes. In addition, seeds from thousands of varieties of wheat, collected from all over the world, are being grown into seedlings and examined for R genes.
603