Concept 29.3: Metabolic Rates Are Affected by Homeostasis and by Regulation and Conformity
The cells inside an animal’s body are bathed with body fluids called tissue fluids or, more formally, interstitial fluids. Suppose we ask, what is the environment of cells inside the body? The tissue fluids, which exist around and between cells, are that environment. For this reason, an animal’s tissue fluids are sometimes called the animal’s internal environment, whereas the outside world is the animal’s external environment.
To see this distinction more clearly, suppose you are standing outside on a cold, dry day. Your external environment will have a low temperature and little moisture. However, inside your body, your body fluids will be warm and wet. The environment of your cells—your body’s internal environment—will therefore be warm and wet.
Animals are classed as regulators and conformers
What happens inside an animal when its external environment changes? A discussion of temperature provides an excellent starting point for understanding the answer.
610
Some types of animals, such as mammals and birds, maintain a constant internal temperature, much in the way that a house with a furnace and an air conditioner maintains a steady inside temperature. Humans, for example, maintain a constant internal temperature of about 37°C (98.6°F). In these types of animals, if the temperature of the external environment changes, the internal temperature does not change. Such animals are called regulators and are said to exhibit regulation. By definition, regulation occurs when the internal environment stays constant even as the external environment changes (FIGURE 29.4A).
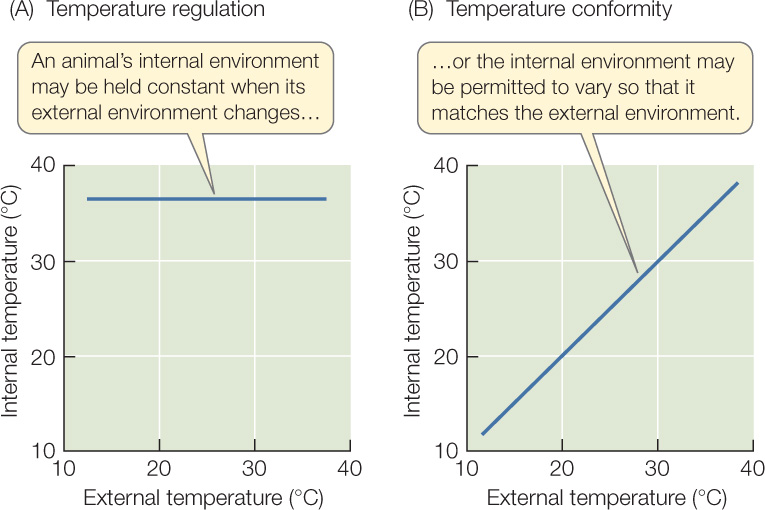
In other types of animals, such as fish and frogs, if the temperature of the external environment changes, the internal temperature changes to match the new external temperature. These animals are called conformers and are said to exhibit conformity. By definition, conformity occurs when the internal environment varies so that it always matches the external environment (FIGURE 29.4B).
The concepts of regulation and conformity can be applied to all characteristics of an animal’s internal and external environments, not just temperature. Moreover, an animal that conforms in some characteristics may regulate in others. We’ve seen, for example, that fish are conformers in regard to temperature. They are regulators, however, in terms of the concentration of sodium (Na+) ions, symbolized [Na+], in their tissue fluids. If the sodium concentration of the external water changes, a fish keeps a constant [Na+] in its tissue fluids, even though it lets its internal temperature change when the external temperature changes.
Regulation is more expensive than conformity
To think further about regulation and conformity, let’s contrast a modern house with a simple log cabin without a stove or furnace. In terms of temperature, when we are inside the modern house, we are always comfortable. This happy state of affairs changes if we move to the cabin. In hot weather, we sometimes boil. In cold weather, we sometimes freeze. We also realize, however, that the cabin is not entirely bad. We have no fuel costs for the cabin, whereas we pay steep utility bills for the house. This contrast helps us understand the advantages and disadvantages of regulation and conformity in animals.
Regulation (such as we have in the modern house) provides stability but is energetically expensive. If an animal maintains a constant internal environment, its body cells enjoy constancy in their surroundings. This is the advantage of regulation. However, the animal must expend energy to maintain a constant internal environment when the outside environment varies. Regulation tends to increase an animal’s metabolic rate. This is the disadvantage of regulation.
Conformity (such as we experience in the log cabin) does not provide stability but is energetically cheap. If an animal simply lets its internal environment change to match its external environment, the animal need invest no energy in the process. However, its cells must cope with considerable variation in their immediate surroundings.
Homeostasis is a key organizing concept
As the nineteenth century ended and the twentieth century began, physiologists were studying humans and “model” nonhuman animals such as dogs used in biomedical research. These investigators gradually realized for the first time that when people are healthy, their blood and tissue fluids are remarkably consistent in their characteristics from time to time. The investigators often focused their studies on the blood plasma, which is the blood solution in which the red blood cells and other blood cells are suspended. The investigators discovered that the concentration of glucose (“blood sugar”) in a healthy person’s blood plasma and tissue fluids is almost constant, whether a meal has just been eaten or not. They likewise discovered that temperature, pH, and the concentrations of major ions are approximately constant in the blood plasma and tissue fluids.
A famous early-twentieth-century physiologist, Walter Cannon, coined the term homeostasis to refer to this stability of the internal environment and the mechanisms that maintain the stability. To this day, homeostasis is an exceedingly important organizing concept in medicine, veterinary medicine, and many other fields of biology. The meaning of homeostasis has shifted a little over time; the term is sometimes used today to refer to any sort of stability or constancy in an animal.
Homeostasis, you will recognize, is a very similar concept to regulation. Homeostasis offers advantages, but as you can guess, it is energetically expensive. Accordingly, when we consider the entire animal kingdom, we find that homeostasis is not universal. Animals vary in their degrees of homeostasis.
Animals are classed as homeotherms or poikilotherms based on their thermal relationships with their external environment
Let’s focus in greater detail on the temperature relationships of animals. These relationships have major effects on animal metabolic rates, as well as being important in many other ways.
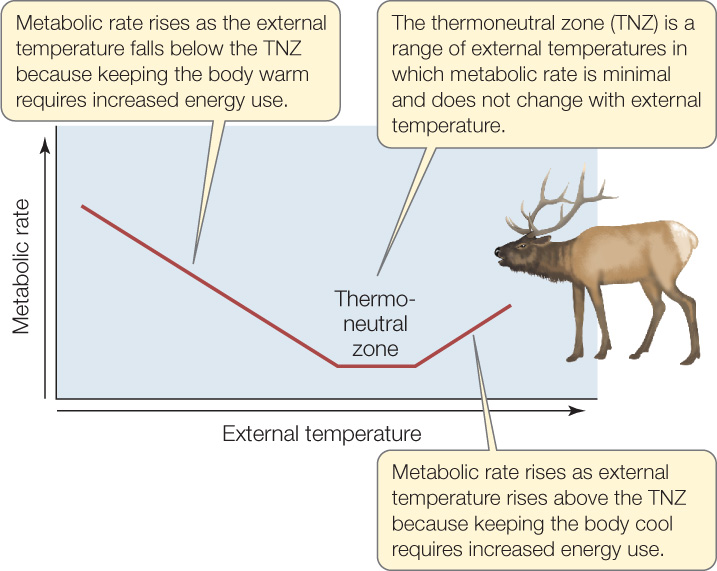
Animals that maintain an approximately constant internal body temperature—such as mammals and birds—are said to exhibit thermoregulation and are called homeotherms. In most homeotherms, if a resting individual is exposed to a variety of external temperatures, its body temperature remains constant but its metabolic rate varies with external temperature as seen in FIGURE 29.5. Over a range of relatively mild external temperatures, called the thermoneutral zone (TNZ), the animal’s metabolic rate is constant.
Go to ACTIVITY 29.1 Thermoregulation
PoL2e.com/ac29.1
611
At temperatures below the TNZ, a homeotherm’s metabolic rate increases as the external temperature falls. This happens for the same reason that the furnace in a house burns fuel faster as the weather outdoors becomes colder. A homeotherm must progressively increase its rate of metabolic heat production as the external temperature becomes progressively colder, in order to offset its progressively increasing rate of heat loss.
At temperatures above the TNZ, a homeotherm’s metabolic rate increases as the external temperature rises. This happens for the same reason that the air conditioner in a house works harder as the weather outdoors becomes hotter. As the animal’s external temperature becomes progressively warmer, the animal must work progressively harder to keep its body temperature from rising. Its metabolic rate reflects this harder work by rising.
Most animals are not homeotherms, and their relationship between metabolic rate and temperature is different than that of homeotherms. Fish, frogs, sea stars, crayfish, and many other kinds of animals permit their body temperature to match their external temperature. Such animals are called poikilotherms or ectotherms. These two names reflect two complementary characteristics of these animals: The animals have body temperatures that are variable and that are determined by the external temperature. The prefix poikilo means “variable,” and ecto means “external.” The name poikilotherm emphasizes the variability of body temperature in these animals, and the name ectotherm emphasizes the fact that their external temperature determines their body temperature. In this book we favor the term poikilotherm.
An important question about poikilotherms is how high their body temperature can rise—and how low it can fall. Poikilotherms vary widely in their body temperature limits:

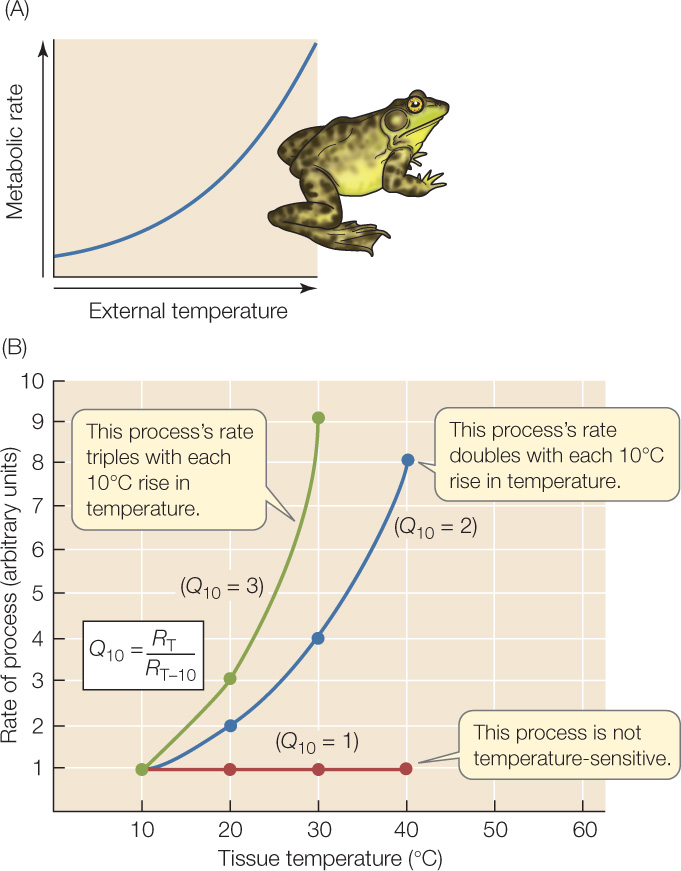
When the external temperature of a poikilotherm rises or falls, the animal’s metabolic rate varies approximately exponentially (FIGURE 29.6A). The reason for this is that the animal’s tissues become cooler or warmer as the external temperature varies. The tissues are cold in a cold environment and warm in a warm environment. Low tissue temperatures slow both biochemical processes (such as enzyme-catalyzed reactions; see Figure 3.22) and biophysical processes (such as diffusion), thereby slowing metabolism. High tissue temperatures do the opposite.
612
In a tissue, we can describe the temperature sensitivity of a reaction or process in numerical terms with Q10, a factor that is easy to calculate. We simply divide the rate of the reaction or process at any particular tissue temperature, RT, by the rate of that reaction or process at a tissue temperature 10°C lower, RT−10:
Q10 = RT/RT−10
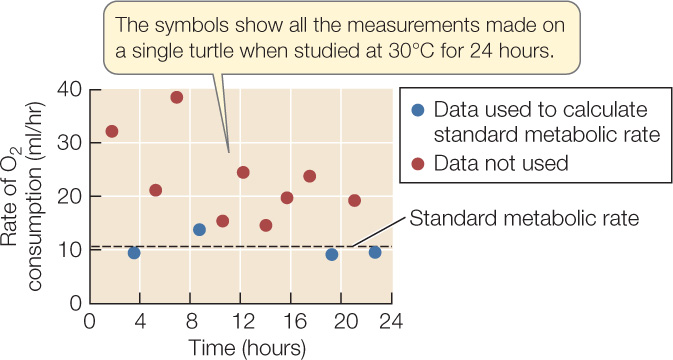
If a reaction or process is not sensitive to tissue temperature, it has a Q10 of 1. Most biological Q10 values are between 2 and 3 (FIGURE 29.6B). A Q10 of 2 means the rate of a reaction or process doubles as tissue temperature increases by 10°C; a Q10 of 3 means the rate triples. FIGURE 29.7 uses the Q10 concept to help interpret how mud turtles respond to changes in temperature.
Investigation
HYPOTHESIS
A mud turtle’s standard rate of metabolism is influenced by its body temperature.
METHOD
- Acclimate a group of wild-caught turtles to an external temperature of 30°C, by letting them live at that temperature for a long time. Deprive them of food so no food processing (which can increase metabolic rate) will occur during the experiment.
- Place each turtle in a sealed chamber connected to a device that measures O2 consumption for 24 hours. Periodically measure the rate of O2 consumption inside the chamber.
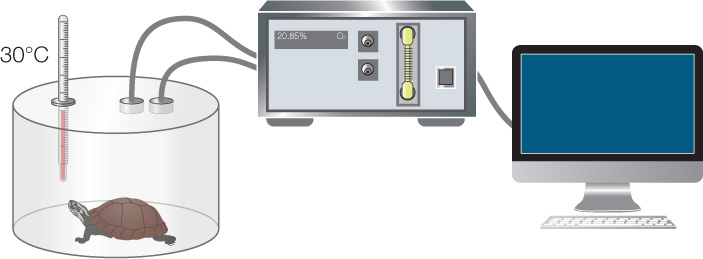 Figure 29.7: The Metabolic Rate of a Poikilotherm Slows Down as Temperature Decreases Jacqueline Litzgus and William Hopkins measured the standard metabolic rate of the mud turtle Kinosternon subrubrum at varying body temperatures.a They showed that temperature has a significant effect on standard metabolic rate in this species. The standard metabolic rate of a poikilotherm is its metabolic rate when resting and fasting. It can be measured at any body temperature of interest. (Poikilotherms do not have thermoneutral zones, explaining why the BMR concept does not apply to them.)
Figure 29.7: The Metabolic Rate of a Poikilotherm Slows Down as Temperature Decreases Jacqueline Litzgus and William Hopkins measured the standard metabolic rate of the mud turtle Kinosternon subrubrum at varying body temperatures.a They showed that temperature has a significant effect on standard metabolic rate in this species. The standard metabolic rate of a poikilotherm is its metabolic rate when resting and fasting. It can be measured at any body temperature of interest. (Poikilotherms do not have thermoneutral zones, explaining why the BMR concept does not apply to them.) - Repeat the acclimation and measurement processes at 20°C.
- From the rates of O2 consumption, calculate the average standard metabolic rate at each temperature.
RESULTS
The average standard metabolic rate at 20°C was 2.10 ml O2 consumed/hour. The average standard metabolic rate at 30°C was 9.25 ml O2 consumed/hour.
CONCLUSION
In turtles, which are poikilotherms, a lower external temperature results in a lower standard metabolic rate.
ANALYZE THE DATA
- Calculate the value of Q10 for the mud turtle’s standard metabolic rate. Based on the Q10 that you calculate, what would you expect the standard metabolic rate (expressed as rate of O2 consumption) to be at 10°C?
- The graph shows data for a single turtle recorded at 30°C. In calculating the turtle’s standard metabolic rate, the researchers chose to use only the lowest 30% of O2 consumption rates recorded. Why might the researchers have made this decision?
Go to LaunchPad for discussion and relevant links for all INVESTIGATION figures.
aJ. D. Litzgus and W. A. Hopkins. 2003. Journal of Thermal Biology 28: 595–600.
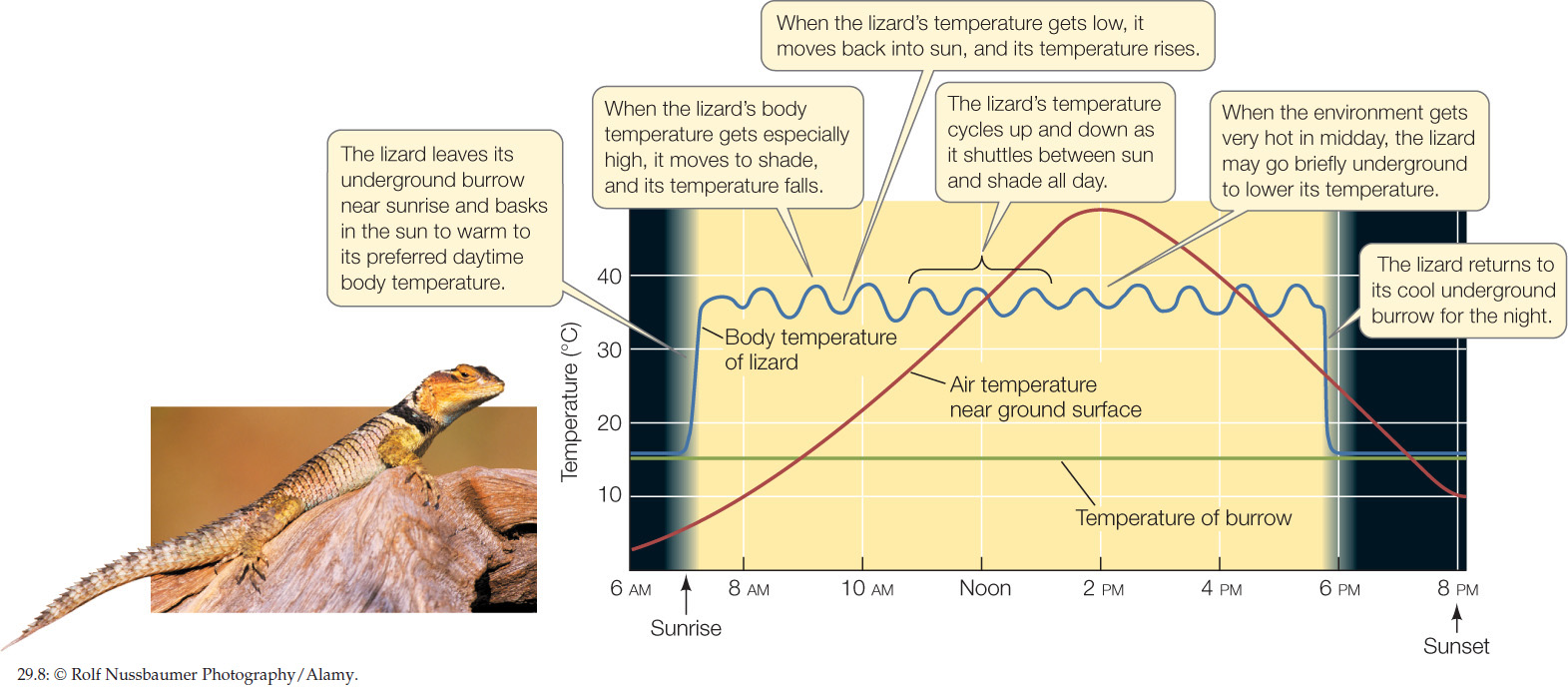
Poikilotherms often exert some control over their tissue temperatures by means of behavior. Because a poikilotherm’s body temperature is determined by external temperature, the animal can adjust its body temperature by positioning itself in suitable environments (FIGURE 29.8). For example, if a poikilotherm’s body temperature is too low, the animal can raise it by moving to a warmer place in its environment (if a warmer place is available).
Homeothermy is far more costly than poikilothermy
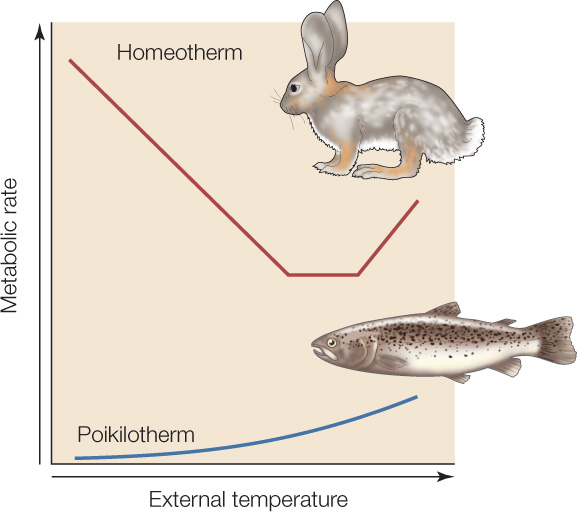
FIGURE 29.9 shows the metabolic-rate-versus-external-temperature relationship for a mammal or bird (homeotherm) on the same plot as that of a fish or lizard (poikilotherm) of the same body size. Notice that at each external temperature, the metabolic rate of the homeotherm is far higher than that of the poikilotherm. Notice especially, toward the left side of the graph, that in cold environments, as the external temperature falls, the metabolic rates of the animals become more different. With a falling external temperature, the homeotherm must make heat faster and faster to keep its tissues warm, whereas the poikilotherm’s tissues become colder, slowing its metabolism.
613
Over the course of an entire year, the average metabolic rates of mammals and birds living in their natural environments are typically 15–30 times higher than the average metabolic rates of fish and lizards in their natural environments (comparing animals of equal body size). This contrast dramatizes the advantages and disadvantages of homeothermy. The cells inside the body of a homeotherm function at approximately one temperature whether the outside environment is cold or warm. The price of this regulation of body temperature is a 15- to 30-fold increase in the animal’s food needs!
LINK
Maintenance of homeostasis can affect life-history tradeoffs—the amount of resources an animal can allocate to reproduction, growth, and defense; see Concept 42.3
Homeotherms have evolved thermoregulatory mechanisms
Mammals, birds, and insects are the major groups of homeothermic animals. Placental mammals have core body temperatures of about 37°C whether they live in cold or warm climates. Birds have higher body temperatures, about 39°C. Among insects, only some exhibit homeothermy. Moreover, the insects that display homeothermy do so just at certain times, such as during flight. Many moths, butterflies, bees, and dragonflies maintain relatively constant, high tissue temperatures in their thorax—the location of their flight muscles—when they are flying. Crickets and katydids are similar when singing. Honeybee hives also display homeothermy. Collectively, the workers in a hive keep the brood (their eggs and larvae) at about 32–36°C even when the outside air temperature is far lower than freezing or as hot as 50°C.

Go to MEDIA CLIP 29.1 Thermoregulation in Animals
PoL2e.com/mc29.1
All homeothermic animals have mechanisms for increasing their metabolic rate—therefore increasing their heat production—when their external environment is cold. In some of these mechanisms, ATP (see Concept 6.1) is the source of energy to make heat. ATP is an energy-shuttle compound in cells. It stores energy when food molecules are broken down. It then can itself be broken down, releasing that energy.
Both mammals and birds shiver to increase heat production. During shivering, skeletal muscles contract in such a way that they produce only subtle, quivering motions. The energy for these contractions comes from the chemical bonds of ATP. As molecules of ATP are used for contraction by the quivering muscles, chemical energy from ATP is converted to heat at a heightened rate.
614
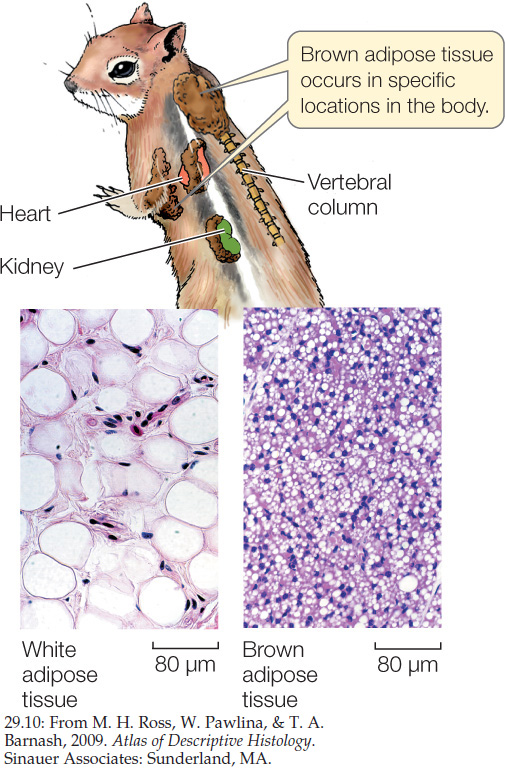
Many mammals, but not birds, are also capable of nonshivering thermogenesis, in which thermogenesis (heat production) occurs without shivering. This process takes place in a specialized type of fatty tissue called brown adipose tissue (BAT) (FIGURE 29.10). The mechanism used by BAT to make heat is uncoupling of oxidative phosphorylation. In this process, no ATP is made. Instead, heat is made directly in the mitochondria because the proton gradient in the mitochondria is short-circuited.
LINK
Uncoupling during oxidative phosphorylation is discussed in Concept 6.2
When insects maintain a warm thorax during preparation for flight or during flight or singing, the flight muscles in their thorax provide the heat needed (FIGURE 29.11). In this case, the heat comes from ATP.
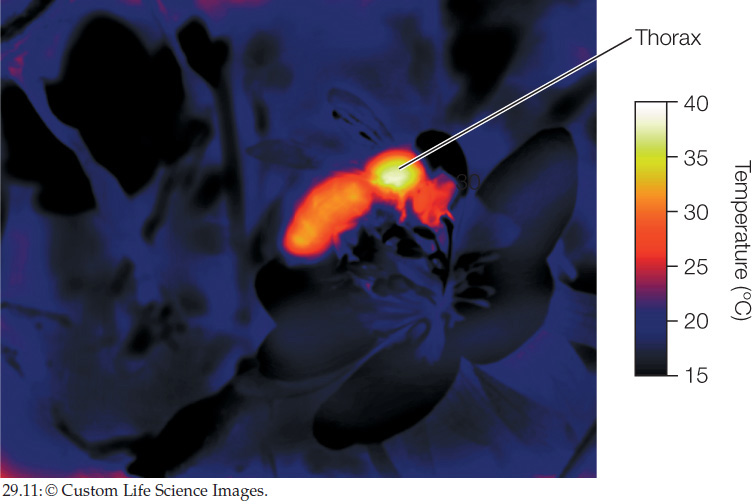
Besides heat production, body insulation is another important characteristic that allows homeotherms to maintain high tissue temperatures in cold environments. Fur and feathers provide high insulation. Heat is also often retained by specialized blood flow patterns (see Concept 32.5).
In hot environments, homeotherms often need to increase their rate of heat loss to keep from overheating. The most common way of doing this is to increase the rate of evaporation of water. When water turns from a liquid to a gas (water vapor), it absorbs a great deal of heat per gram (see Concept 2.2). This heat is removed from the animal when the vapor travels away in the atmosphere. Many large mammals—such as humans, horses, and camels—sweat to increase their rate of evaporation. Dogs pant. Honey bees in a hive spread water that they have collected on interior surfaces of the hive and then fan the moistened surfaces with their wings to speed evaporation.
Hibernation allows mammals to reap the benefits of both regulation and conformity
Mammalian hibernation is remarkable in many ways—one being that hibernators are able to reap the benefits of both regulation and conformity. Most hibernating species are relatively small. They include certain mice, hamsters, ground squirrels, groundhogs, marmots, bats, and small marsupials.
The usual pattern for a hibernator is that the animal functions as a homeotherm—regulating its body temperature near 37°C—during the warm months of the year. During the cold months, however, the animal suspends homeothermy and exhibits temperature conformity. It then allows its body temperature to match the external temperature over a wide range of temperatures, meaning in many cases that the body temperature falls as low as 0–10°C (FIGURE 29.12). Hibernation is the term for this winter period characterized by conformity.
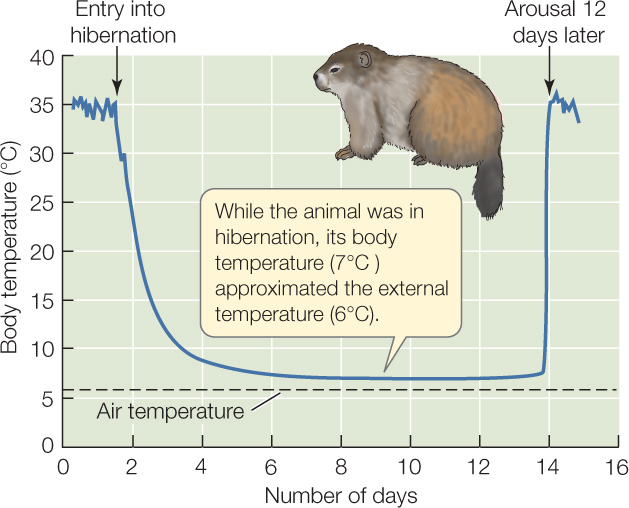
When an animal functions as a temperature regulator, it experiences the advantages of a high, stable body temperature: its cells function in a consistent thermal environment. The animal faces high metabolic costs, however, like other homeotherms. When the animal functions as a temperature conformer, it experiences the advantages of conformity, namely that its metabolic costs are very low. Hibernators essentially switch between two metabolism–temperature curves resembling those in Figure 29.9.
By reducing energy needs, hibernation allows an animal to spend the winter continuously in a relatively protective and safe place, such as an underground burrow. The amount of metabolic fuel the animal requires is reduced to the point that the animal need not roam around its habitat in search of food. Instead, it can live entirely on body fat or on stored food it collected the previous summer.
615
CHECKpoint CONCEPT 29.3
- How would the pH of the interstitial fluids of a healthy, water-dwelling regulator be affected by a moderate increase in the pH of the external environment?
- Would a reaction that proceeds faster as temperature increases have a Q10 value higher or lower than 1?
- What is one advantage and one disadvantage of the conforming lifestyle?
- Does the metabolic rate of a homeotherm increase or decrease as external temperature decreases? Assume the temperature is below the homeotherm’s thermoneutral zone. Would the answer differ for a poikilotherm?
Now that we’ve discussed how animals use energy, let’s consider their body organization and the ways they make ATP.