Concept 29.6: Animal Function Requires Control Mechanisms
The nervous and endocrine systems are the principal systems that control or regulate the ways tissues and organs interact harmoniously throughout an animal’s body. In addition, there are control systems in individual cells that control cellular processes such as biochemical reactions and cell division. The importance of controls is emphasized dramatically when controls fail. Cancer in part reflects a failure of control of cell division. Lethal fevers reflect failure of control of body temperature.
LINK
Cancer and regulation of the cell cycle are discussed in Chapter 7; see Concept 7.3
Homeothermy exemplifies negative-feedback control
In discussing animal control systems, the control of body temperature in humans and other mammals is a helpful example because it takes place in ways analogous to the familiar engineered systems we use to regulate temperature in our homes, as we mentioned earlier. Not all animal control systems operate in the same way as the control of body temperature, but many do.
The four essential elements of a control system are the controlled variable, sensors, effectors, and control mechanism. The controlled variable is the characteristic of an animal that is being controlled—body temperature in this example. Sensors detect the current level of the controlled variable. In body temperature regulation, the sensors are neural structures that detect the existing temperature in various parts of the body (e.g., skin, brain, and scrotum) and send the information to the control mechanism in the brain. Effectors are tissues or organs that can change the level of the controlled variable. Shivering muscles, for example, are effectors because—by generating heat—they can change the body temperature. The control mechanism uses information from the sensors to determine which effectors to activate—and how intensely—to modify the controlled variable. The control mechanism for body temperature is composed of nerve cells located in and near the hypothalamus of the brain.

Go to ANIMATED TUTORIAL 29.1 The Hypothalamus
PoL2e.com/at29.1
In the regulation of mammalian body temperature, as shown in FIGURE 29.16, when the control mechanism receives information on the existing body temperature from the sensors, it compares the current body temperature with a set-point temperature—the temperature that is supposed to prevail, in this case 37°C. The control mechanism then activates effectors to reduce or remove the difference between actual body temperature and the set point. If the body temperature is lower than the set point, the control mechanism activates shivering or nonshivering thermogenesis, resulting in increased heat production, which tends to raise the body temperature until it equals 37°C. If the body temperature is higher than the set point, the control mechanism activates sweating or panting to reduce or remove the difference.
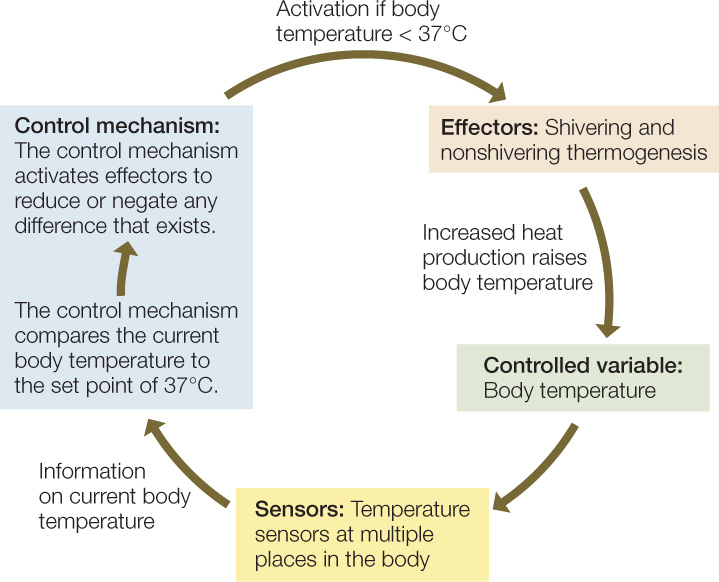
This control mechanism is an example of a negative-feedback system. The process is called “feedback” because after the control mechanism issues commands to effectors, it receives information on how its commands have altered the body temperature (the controlled variable). Information is “fed back” to the control mechanism. The process is described as “negative” because the control mechanism activates effectors in ways that reduce or negate any difference that exists between the body temperature and the set point. Negative feedback is a stabilizing influence and is often involved in maintaining homeostasis. In this case, it keeps the body temperature near 37°C.
620
Positive feedback occurs in some cases
Positive feedback occurs when deviations of a controlled variable from its existing level are increased or amplified by the action of the control mechanism. Positive feedback destabilizes a system. However, destabilization can sometimes be advantageous, provided it is ultimately brought under control. A nerve impulse is an example of positive feedback that lasts for a very brief time and then is terminated (see Figure 34.8).
The mammalian birth process depends on positive feedback. During the birth process, waves of contraction of a mother’s uterine muscles are stimulated by the hormone oxytocin, which is secreted by neurons in the hypothalamus of the brain and released into the blood by the posterior pituitary gland. A control mechanism in the hypothalamus receives information from sensors in the uterus on the force of uterine contractions. When this control mechanism detects an increased force of contraction, it activates increased oxytocin secretion by the neurons in the hypothalamus, which are the effectors in this system. The heightened oxytocin secretion further increases the force of contraction in a self-reinforcing, positive-feedback loop (FIGURE 29.17). Ultimately the force becomes great enough to cause the fetus to move from the uterus into the outside world, ending further stimulus for oxytocin production and uterine contractions.

Biological clocks make important contributions to control
Animals have self-contained, metabolic mechanisms of keeping track of time, known as biological clocks. These mechanisms do not require outside information to function. Instead, they have a self-contained ability to keep time. Biological clocks have several advantages. One of the most important is that they allow animals to anticipate future events, just as our watches let us anticipate when future events will occur.
The early experiments that convinced biologists of the existence of biological clocks focused on activities such as wheel-running in small mammals, such as mice and squirrels. Given a running wheel in a normal cage environment with day and night, nocturnal species of squirrrels tend to run on the wheel each night and to be inactive each day. They alternate between running and not running on a 24-hour cycle (FIGURE 29.18A). Investigators found that if they deprived these squirrels of an external day–night cycle, for example by keeping them in constant darkness, the squirrels still tended to alternate between periods of high and low running activity on an approximately 24-hour cycle (FIGURE 29.18B). This suggested that squirrels can keep time without external timing cues. To make sure that the squirrels were not receiving timing cues from their surroundings, investigators not only deprived the animals of an external day–night cycle but also isolated them from outside temperature variations, outside sounds, and any other outside cues imaginable. The squirrels still alternated between periods of high and low running activity. By now, based on revolutionary discoveries of the past 20 years, the mechanisms of biological clocks have been deciphered. Thus the existence of biological clocks has been confirmed by knowledge of the ways they work.


Go to ANIMATED TUTORIAL 29.2 Circadian Rhythms
PoL2e.com/at29.2
When an animal is prevented from receiving external timing cues, its biological clocks are said to free-run, and under these conditions they do not keep perfect time (see Figure 29.18B). They run fast or slow relative to actual time. Consider a biological clock that, in its free-running state, goes through an entire timing cycle in 23.8 hours. After 1 day of free-running, the clock will be 0.2 hour off, compared with the actual 24-hour solar-day rhythm. After 2 days it will be 0.4 hour off, and after 5 days it will be an entire hour off. In natural environments, this long-term drift is prevented because animals receive external timing cues and their biological clocks do not free-run. Each day, the clocks are reset to be synchronous with actual time—a process termed entrainment. Sunrise and sunset are the most commonly used cues for entrainment. In a natural environment, animals observe sunrise and sunset each day, and they use these indicators to synchronize their biological clocks with the actual 24-hour solar-day rhythm.
Biological clocks that have free-running timing cycles of about 24 hours are called circadian clocks. The word “circadian” means “about a day.” It emphasizes that the self-contained, free-running periodicities of these clocks are not exactly 24 hours. By now we know of the existence of biological clocks that have free-running timing cycles of about 1 year, called circannual clocks. Many animals in the ocean, where there are lunar tides, have circatidal clocks. All require entrainment to keep exact time.
621
Circadian clocks are believed to be virtually universal among animals. In an individual animal, most or all cells have their own circadian clocks. A “master clock” is present in an individual, however, and sends out signals to other cells in the individual’s body, keeping the body as a whole synchronized. In mammals, the master clock consists of nerve cells in parts of the brain called the suprachiasmatic nuclei.
APPLY THE CONCEPT: Animal function requires control mechanisms
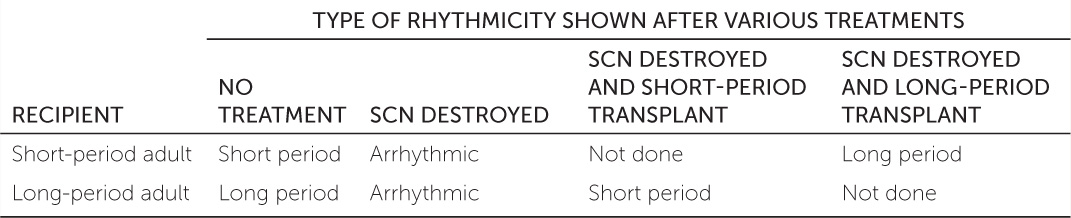
To better understand the physiological mechanism of circadian clocks, Martin Ralph and colleagues used artificial selection to produce two strains of hamsters: one with a short circadian period and one with a long circadian period.a (The circadian period is the length of time between two successive days as timed by the clock.) When the suprachiasmatic nuclei (SCN; parts of the brain involved in controlling circadian rhythms) in some of these adult hamsters were destroyed, the animals became arrhythmic. After several weeks, the scientists transplanted SCN tissue from fetal hamsters into the brains of the adult hamsters with destroyed SCNs. Long-period adult hamsters received tissue from short-period fetuses, and short-period adults received tissue from long-period fetuses. The effects of these treatments on the hamsters’ circadian periods are shown in the table.
- Why did destroying the SCN make the adult hamsters arrhythmic?
- What result would you expect if SCN tissue from a long-period fetal hamster were transplanted into a long-period adult that had a destroyed SCN?
- What do the results of the transplantation experiments tell you about where in the body the circadian rhythm phenotype (long versus short) is expressed?
a M. R. Ralph et al. 1990. Science 247: 975–978.
622
Biological clocks help control many animal functions. Circadian clocks can give animals an internal sense of when to wake up each day. They help control when animals are active and at rest. They also help control when the digestive system is most able to digest foods each day and when detoxification systems are most able to counteract toxic chemicals. Circannual clocks are known to control annual hibernation rhythms in some species of mammals, and they control annual reproductive rhythms in some animals, such as sheep.
LINK
Plants also have circadian clocks, which are entrained by light cues; see Concept 26.4
CHECKpoint CONCEPT 29.6
- Imagine a house with an air conditioner, furnace, and electric thermostat consisting of a thermometer and simple computer. For this control system, identify the controlled variable, sensor(s), effector(s), and control mechanism. Draw a negative-feedback loop for cooling a house using these components.
- What is the likely fate of a biological clock that is denied any chance for entrainment?
- Does negative feedback always have a diminishing effect on the controlled variable? That is, because it’s “negative,” does it always make the level of the controlled variable very low?
Question 29.2
Why can we use a person’s rate of O2 consumption to measure the rate at which he or she consumes energy to run?
ANSWER To be able to do muscular work, muscle cells need ATP, which they must make themselves (Concept 29.4). As food molecules are broken down in muscle cells to provide energy to synthesize the ATP needed to power the work of muscles, O2 is required in proportion to the energy released from the food molecules (Concept 29.2). Thus, logically, O2 is required in proportion to the ATP that is made and broken down in metabolism. By measuring the rate at which O2 is consumed, we can calculate the rate of breakdown of food molecules to release energy, the rate of ATP synthesis from that energy, and the rate of heat production as muscles liberate energy from ATP and use it to contract, converting it to heat.
