Concept 30.4: The Vertebrate Digestive System Is a Tubular Gut with Accessory Glands
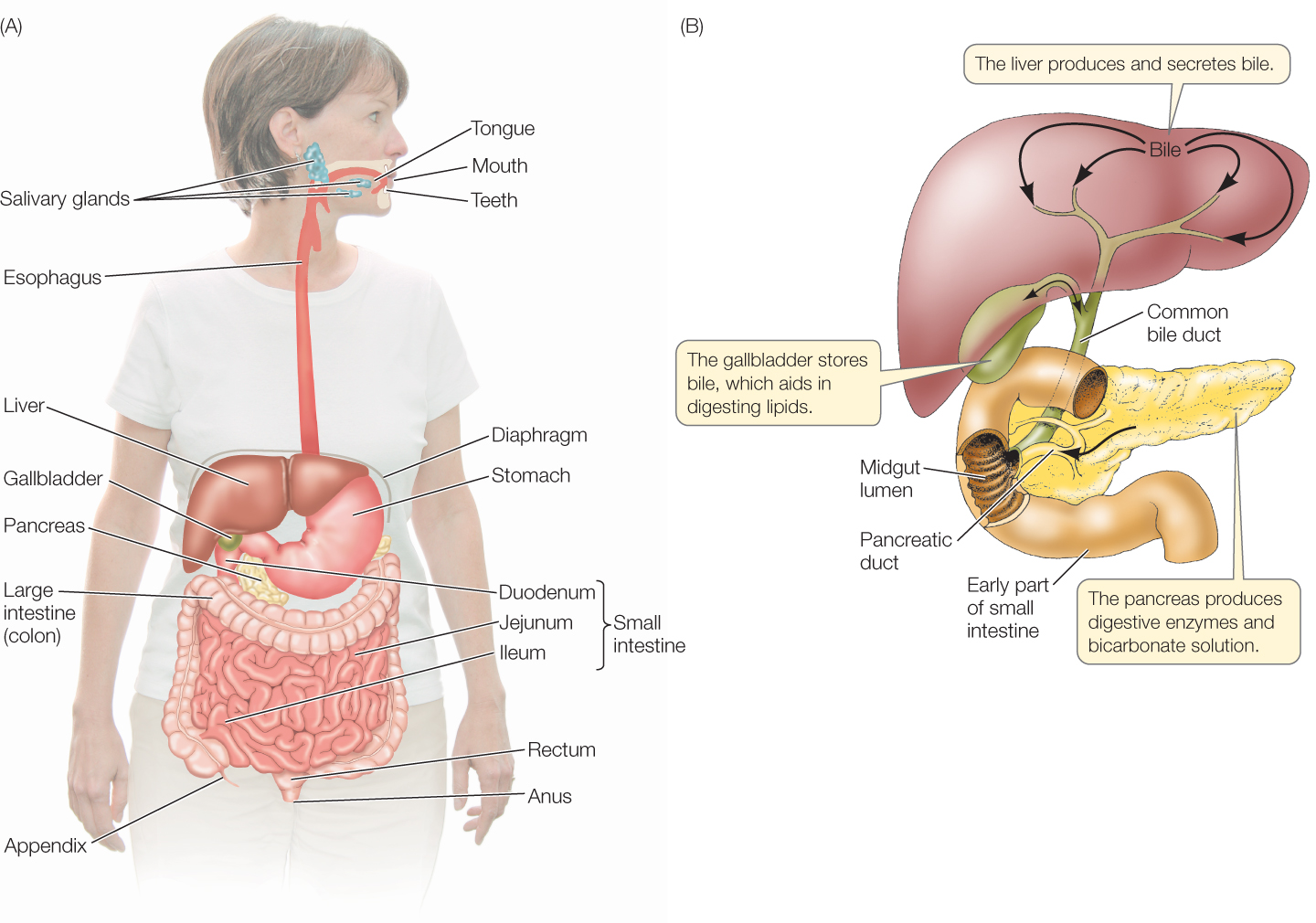
The vertebrate digestive system consists of a tube, extending from mouth to anus and called the gut, that receives secretions from certain nearby glands, most notably the liver and pancreas (FIGURE 30.11; also see the bottom panel of Figure 30.8). The tissues of the gut wall are arranged in concentric layers (FIGURE 30.12). Surrounding the gut lumen is the gut epithelium, a functionally dynamic cell layer. Most cells in the epithelium are digestive–absorptive cells, which produce distinctive proteins, such as digestive enzymes, that function in digestion and absorption. In addition to the dominant digestive–absorptive cells, the gut epithelium also contains scattered mucus-secreting cells, plus scattered endocrine cells that secrete hormones involved in controlling the digestive tract (as you will see in Concept 30.5). Surrounding the epithelium and its underlying connective tissue (together called the mucosa) is a tissue layer called the submucosa, which contains blood and lymph vessels that carry absorbed nutrients to the rest of the body. The submucosa also contains a network of neurons that help control the secretory functions of the gut.
Go to ACTIVITY 30.3 The Human Digestive System
PoL2e.com/ac30.3
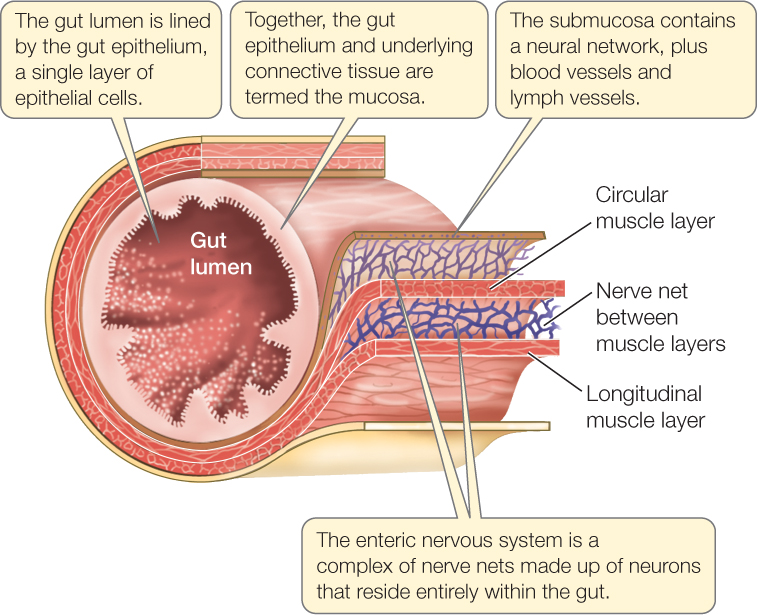
Two layers of smooth muscle surround the submucosa. The inner layer, composed of circular muscles, encircles the gut and constricts it when it contracts. The outer layer, composed of longitudinal muscles, shortens the gut when it contracts. Between the two muscle layers is a second network of neurons, which coordinates contractions of the muscle layers so that food materials in the gut are pushed along by waves of contraction called peristalsis or, at times, are pushed back and forth. The passage of materials through the gut is further controlled by sphincter muscles located at several places, such as the entrances to the stomach and small intestine. Sphincter muscles encircle the gut, and they contract so tightly that they seal off one part of the gut from another except when they are opened briefly in a controlled manner to let food materials pass through.
The neuron networks in the gut wall make up the enteric nervous system. This system is part of the autonomic nervous system, which controls many body functions without any need of conscious input and, in general, without voluntary control (see Concept 34.5).
An interesting aspect of the open central cavity of the gut tube, the gut lumen, is that it is anoxic, meaning it is devoid of O2. Tapeworms and other gut parasites thus must have the ability to live without O2, a very unusual ability for animals.
Several classes of digestive enzymes take part in digestion
Like most enzymes, the digestive enzymes are proteins. They break chemical bonds by hydrolysis, a reaction that splits a bond by adding a water molecule (see Figure 2.8). The digestive enzymes are classified in two ways. One is based on the type of food molecules the enzymes digest. Lipases hydrolyze bonds of lipids; proteases and peptidases hydrolyze the bonds between adjacent amino acids in proteins; and carbohydrases hydrolyze the bonds between adjacent sugar units of carbohydrates. Besides these general names, specific names are used for specific enzyme proteins.
636
Digestive enzymes are also classed by where they act:
- Intraluminal enzymes are secreted into the gut lumen and digest food molecules in the lumen.
- Membrane-associated enzymes are attached to the epithelial cells lining the gut lumen. They act when food molecules in the lumen come in contact with the gut epithelium.
- Intracellular enzymes are inside some of the gut epithelial cells and are involved in protein digestion.
Digestive enzymes typically act in sequences. Food proteins, for example, are first attacked by protein-digesting enzymes in the stomach that break the protein molecules into relatively large pieces. Other protein-digesting enzymes in the intestines then break these pieces into smaller pieces.
Processing of food starts in the foregut
The digestive tract begins with the foregut, which consists of the mouth, esophagus, and stomach—and in some animals, an expanded food-storage chamber (crop) or food-grinding chamber (gizzard) ahead of the stomach. When most vertebrates eat, they simply seize food with their mouth, then swallow it whole. Fish, amphibians, birds, and non-avian reptiles eat this way. Mammals, however, chew their food by using their teeth and jaw muscles, and as they chew, they start the process of carbohydrate digestion by secreting saliva from their salivary glands into their mouth cavity. Mammalian saliva contains a starch-digesting enzyme, amylase.
637

Go to MEDIA CLIP 30.1 Following Food from Mouth to Gut
PoL2e.com/mc30.1
The esophagus typically serves simply as a passageway for food to reach the stomach from the mouth. Each time we swallow, a sphincter at the start of our esophagus opens briefly (under control of the enteric nervous system) to let food pass from our mouth into our esophagus. Then, after peristalsis has moved the food from the start of the esophagus to the end, the sphincter at the entrance to the stomach briefly opens to permit the food to enter the stomach.
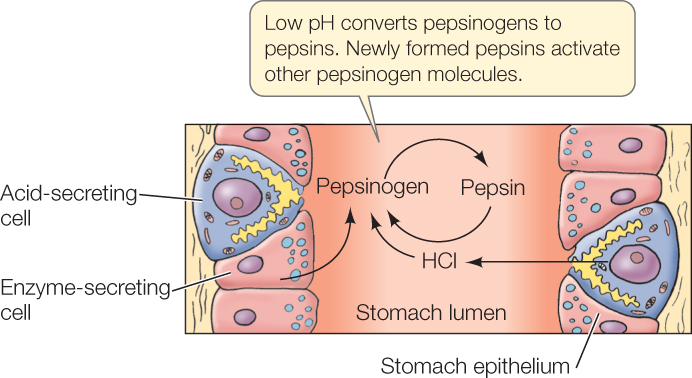
The stomach is an expanded part of the foregut that, in most vertebrates, performs four major functions. First, the stomach serves as a storage chamber for newly ingested food. It can hold a large meal that is awaiting processing. Second, the stomach secretes hydrochloric acid (HCl), which mixes with food in the stomach lumen, helping cause food disintegration. When you think about the pHs that are sometimes achieved in the stomach, you will probably be amazed that living cells are capable of secreting the acid needed. In humans and dogs, the contents of the stomach lumen during meal processing can reach a pH of 0.8—the same as that of battery acid and ten times more acidic than lemon juice! Acid-secreting cells in the stomach epithelium secrete protons (H+ ions) into the lumen, using energy from ATP, to achieve this astounding pH. The third major function of the stomach is to begin protein digestion. Certain cells in the stomach epithelium secrete protein-digesting enzymes called pepsins into the stomach lumen, where these enzymes start to break up proteins. Finally, the stomach mechanically squeezes food and mixes it with the acid and digestive enzymes. This is achieved by contractions of the smooth muscles in the stomach wall.
A fascinating challenge that had to be met during evolution arises from the fact that protein-digesting enzymes could potentially digest the very cells that synthesize them, killing the cells! The evolutionary solution to this challenge is that these enzymes are synthesized and secreted in inactive forms. Stomach cells synthesize and secrete pepsins as inactive pepsinogens, for example. Only after the pepsinogens arrive in the stomach lumen are they activated (by exposure to HCl) to form pepsins:
As we have noted, some vertebrates have evolved a fermentation vat for microbial symbionts in specialized, nonacidic parts of the foregut. All such animals are classified as foregut fermenters. The ruminants are one such group (see Figure 30.7). Kangaroos, hippos, sloths, colobus monkeys, and a few ground-dwelling, herbivorous birds are also foregut fermenters.
Food processing continues in the midgut and hindgut
After leaving the stomach, food travels through the midgut and hindgut, which are the first and second parts of the intestines. In humans and other mammals, the two parts differ in diameter: the midgut has a smaller diameter and is termed the small intestine; the hindgut has a larger diameter and is called the large intestine. The distinction between “small” and “large” does not apply in many vertebrates, however, because the two parts—although they differ in function—are the same in diameter. This is often the case in fish, for instance.
Fermentation vats for heterotrophic microbial symbionts have evolved as parts of the midgut or hindgut in some vertebrates. These animals, like the foregut fermenters, are all herbivores. Some fish, including catfish, carp, and tilapia, are midgut fermenters. Horses, zebras, rabbits (see Figure 30.8), rhinos, and apes are hindgut fermenters. So are geese, chickens, and ostriches. All profit from being able to ferment cellulose and hemicelluloses, greatly enhancing the energy value of plant foods.
The midgut is the principal site of digestion and absorption
The midgut is the principal site of digestion of all three major food categories: proteins, carbohydrates, and lipids. It is also typically the principal site of nutrient absorption. The products of digestion of all three categories of food are absorbed in the midgut, as are vitamins, minerals, and water.
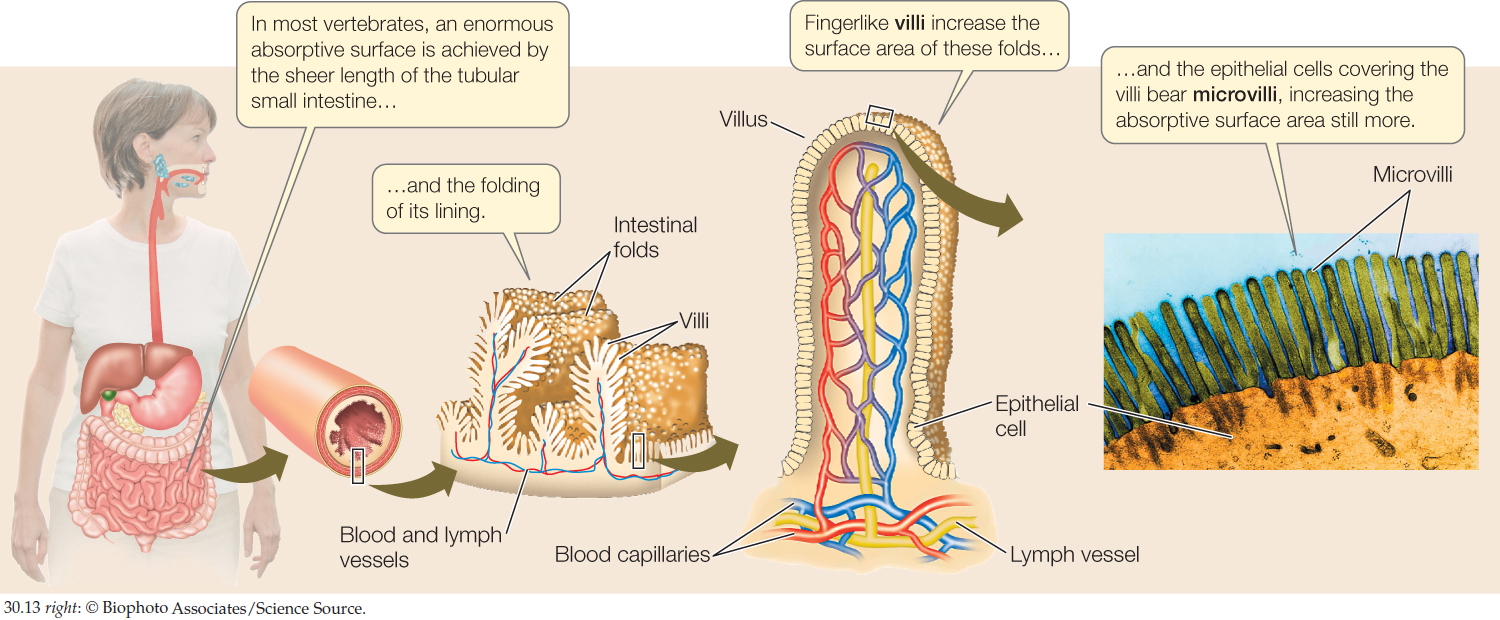
In many vertebrates the inner midgut wall is highly folded, and the folds bear fingerlike projections called villi (singular villus) (FIGURE 30.13). In turn, each cell of the gut epithelium bears many microscopic projections called microvilli. Together, the midgut wall foldings, the villi, and the microvilli vastly enlarge the area of the luminal surface of the midgut epithelium (luminal surface = the surface facing toward the gut lumen). This huge surface area is of great functional importance for two reasons. First, many membrane-associated digestive enzymes are attached to the luminal surface of the gut epithelium. Second, absorption occurs across this surface.
Material from the stomach enters the midgut in a regulated manner. A sphincter muscle, the pyloric sphincter, seals off the stomach from the midgut when it is contracted. Under control of hormones and the enteric nervous system, this sphincter opens briefly to allow controlled, small squirts of digesting material to enter the midgut from the stomach. This measured introduction of material into the midgut ensures that the midgut digestive and absorptive processes are not overwhelmed and can keep up.
Both the pancreas and the liver are connected by ducts to the early part of the midgut (see Figure 30.11B). Digestive juices secreted by the pancreas and emulsifying agents secreted by the liver flow into the midgut lumen via these ducts. The pancreatic digestive juices contain a host of digestive enzymes, including ones that hydrolyze proteins, carbohydrates, and lipids. They also contain bicarbonate ions (HCO3−) that neutralize the highly acidic pH of the material that enters the midgut from the stomach. The pancreas synthesizes and secretes protein-digesting enzymes as inactive molecules termed proenzymes. Only after they reach the midgut lumen are they enzymatically activated to hydrolyze food proteins.
638
Large food molecules are broken down into their smallest molecular subunits during digestion in the midgut. Proteins are broken down into amino acids; carbohydrates into simple monosaccharides (see Concept 2.3) such as glucose, galactose, and fructose; and lipids into fatty acids and glycerol (glycerin) (see Concept 2.4). These small molecules are then absorbed.
Let’s now look briefly at the digestion and absorption of each of the major food categories. Recall that protein digestion starts in the stomach. It continues in the midgut lumen, where the products of stomach digestion are broken down further by protein-digesting enzymes made by the pancreas and gut epithelium (see Figure 30.9). These processes produce both single amino acids and short chains of two or three amino acids, which are absorbed into midgut epithelial cells. Inside these cells, the short chains are further hydrolyzed, so that the final product of protein digestion consists of just single amino acids. These are then transported into the blood vessels in the submuscosa, completing absorption.
In mammals, carbohydrate digestion starts in the mouth with the action of saliva enzymes that help break down starches. It is completed in the midgut. Often the final step in carbohydrate digestion in the midgut is the hydrolysis of a disaccharide—consisting of two linked simple sugar molecules—into two monosaccharide (simple sugar) molecules. The enzymes responsible are called disaccharidases. (Two of the midgut disaccharidases—lactase and trehalase—have been discussed earlier in this chapter.) The monosaccharides are then absorbed across the cells of the midgut epithelium to enter the submucosal blood vessels.
Lipid digestion starts and ends primarily in the midgut. Lipids present a special challenge for digestion because they are not water-soluble. The liver meets this challenge by secreting bile into the midgut. Bile salts and other bile components act as detergents, emulsifying food lipids in the midgut lumen into great numbers of tiny droplets. The lipids then have a high surface area, allowing lipid-digesting enzymes to break the lipids into fatty acids and glycerol. The bile salts then continue to associate with the fatty acids, keeping them in solution until the fatty acids are absorbed into midgut epithelial cells. An interesting characteristic of bile salts is that they are not modified by this process, and in fact are reabsorbed and then reused in processing one meal after another. After fatty acids and glycerol are absorbed into midgut epithelial cells, they are reassembled into simple lipid molecules, which form aggregates with proteins synthesized by the cells. These lipoprotein aggregates—in which the lipids are emulsified by the proteins—then pass into lymph vessels in the submucosa, and later they enter the bloodstream.
The cellular mechanisms by which digestive products enter and leave the gut epithelial cells during absorption include diffusion, facilitated diffusion, and secondary active transport. Of these, the most important is secondary active transport (see Concept 5.3), which requires ATP that the midgut epithelial cells synthesize.
LINK
The various mechanisms for moving molecules across membranes are discussed in Concept 5.2 and Concept 5.3
The hindgut reabsorbs water and salts
The hindgut—called the large intestine or colon in mammals—has two main functions. First, it stores indigestible wastes, in the form of accumulated feces, between defecations. The feces are stored in the final portion of the hindgut, the rectum, until eliminated.
Second, the hindgut completes the reabsorption of needed water and salts from the gut contents prior to elimination of the feces. This may sound like a humble function, but it is vitally important. In humans, about 7 liters of a watery solution containing Na+, Cl−, and other ions are secreted from the blood into the gut per day to facilitate processing of food. This large volume needs to be returned to the blood. Diseases such as cholera and dysentery can block reabsorption of water and salts by the hindgut, and in such cases a person can rapidly lose such massive quantities of water and salts through diarrhea that death quickly results.
639
CHECKpoint CONCEPT 30.4
- Digestive enzymes fall into three classes based on where they act. What are these three classes, and where do they act?
- Using correct terminology, explain how the cells that synthesize protein-digesting enzymes avoid being digested by the enzymes.
- Catfish, carp, and tilapia are some of the fish that are most useful for producing fish biomass for human consumption in fish farming (aquaculture). One reason for this is that they can be fed plant foods—in contrast, for example, to salmon, which are carnivores and therefore must be fed fish. In this context, why is it important that catfish, carp, and tilapia are fermenters?
We have seen how the vertebrate digestive system digests and absorbs a single meal. Finally we will look more closely at nutrition from meal to meal.