Concept 30.5: The Processing of Meals Is Regulated
Most animals do not eat continuously. Instead, they alternate between an absorptive state (food in the gut) and a postabsorptive state (no food in the gut). This alternation poses two important challenges. First, for blood homeostasis to be maintained, the concentrations of glucose and other nutrient molecules in the blood plasma must be stabilized, so they do not skyrocket during meals and plummet between meals. Second, nutrients must be stored while they are abundant during the absorptive state and then released from those stores between meals.
Hormones help regulate appetite and the processing of a meal
Vertebrates have more than ten different types of scattered endocrine (hormone-producing) cells in the gut epithelium. The endocrine cells in one region secrete different hormones than the endocrine cells in other regions. This means that location-specific endocrine signals can be sent between gut regions to help coordinate the processing of a meal, complementing the coordination provided by the enteric nervous system.
When an animal eats and food arrives in its stomach, endocrine cells in the stomach epithelium secrete the hormone gastrin into the blood. Gastrin then travels in the blood to all the other stomach epithelial cells, some of which are specialized to synthesize acid (HCl) or pepsins. Gastrin stimulates these cells to secrete acid and pepsins, which help break down the food (FIGURE 30.14).
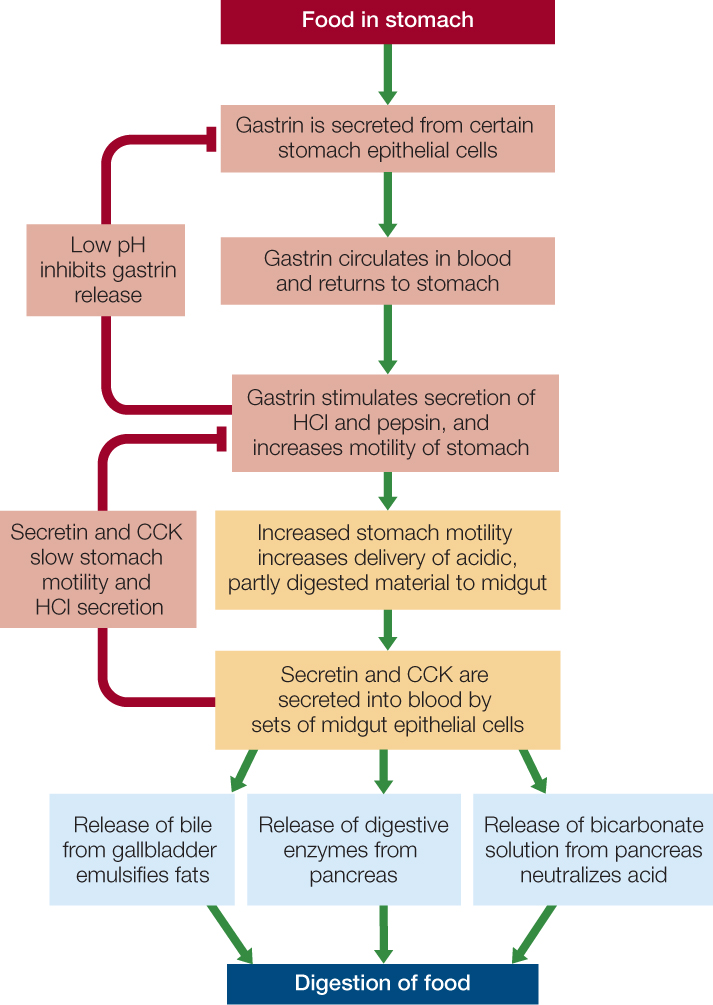
The entry of acidified, partly digested food material into the midgut initiates several important endocrine controls. Recall that the material is released into the midgut in small bursts. Each burst stimulates two distinct sets of endocrine cells in the midgut epithelium (actually there are more than two sets, but we will discuss two here). One set releases secretin (the first hormone ever identified) into the blood, and the other releases cholecystokinin (CCK). When these two hormones reach the stomach, they tend to inhibit stomach acid secretion and muscle contraction (see Figure 30.14). This inhibition is maintained until the midgut has processed the current burst of material from the stomach. The two hormones also affect the pancreas and liver. Secretin stimulates the pancreas to secrete bicarbonate, which flows in the pancreatic juice to the midgut, where it neutralizes the acidity of the recently arrived stomach material. Secretin and CCK, acting synergistically, also stimulate the pancreas to secrete its digestive enzymes—and the liver to secrete bile—into the midgut.
Appetite is also controlled in part by hormones. For example, when the stomach is empty, it secretes ghrelin (pronounced gre-lin), a hormone that stimulates appetite. Another hormone, leptin (named after a Greek word for “thin”), provides information to the brain to suppress appetite. Leptin is secreted by fat cells. High leptin levels indicate that there are many fat cells, and thus there is not a great need to eat. Leptin was discovered, and its properties are still being explored, through studies of mutant mice (FIGURE 30.15).
Investigation
HYPOTHESIS
Mice that cannot produce the protein leptin will not become obese if they are able to obtain leptin from an outside source.
METHOD
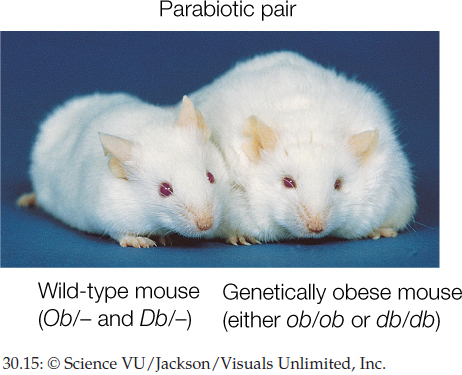
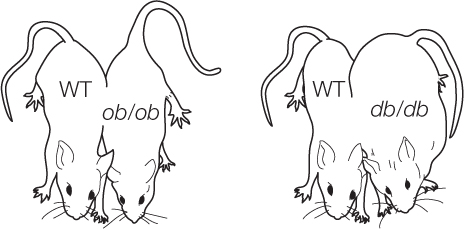
- Create two strains of genetically obese laboratory mice, one of which lacks functional leptin (genotype ob/ob) and one which lacks the receptor for leptin (genotype db/db).
- Create parabiotic pairs by surgically joining the circulatory systems of a non-obese (wild-type) mouse with a partner from one of the obese strains.
- Allow mice to feed at will.
RESULTS
Parabiotic ob/ob mice obtain leptin from the wild-type partner and lose fat. Parabiotic db/db mice remain obese because they lack the leptin receptor and thus the leptin they obtain from their partner has no effect.
CONCLUSION
The protein leptin is a satiety signal that acts to prevent overeating and resultant obesity.
ANALYZE THE DATA
In a further experiment, leptin was purified and injected into ob/ob and Ob/Ob (wild-type) mice daily. The data in the table were collected before the injections began (baseline) and 10 days later.
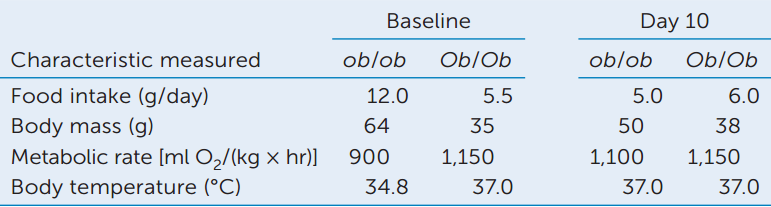
- Do the data support the hypothesis that leptin is a satiety signal? Why or why not?
- Discuss what factors might explain the loss of body mass in the ob/ob mice.
Go to LaunchPad for discussion and relevant links for all INVESTIGATION figures.
aD. L. Coleman and K. P. Hummel. 1969. American Journal of Physiology 217: 1298–1304.
D. L. Coleman. 1973. Diabetologia 9: 294–297.
640

Go to ANIMATED TUTORIAL 30.1 Parabiotic Mice Simulation
PoL2e.com/at30.1
Insulin and glucagon regulate processing of absorbed food materials from meal to meal
Insulin and glucagon are two of the most important hormones that regulate blood nutrient levels—and the storage and release of nutrients in the body—as animals switch back and forth between the absorptive and postabsorptive states. Both hormones originate in the pancreas. The pancreas is composed of two major parts. The cells in one part perform the exocrine functions we have already discussed—secreting digestive enzymes and bicarbonate into the pancreatic ducts to be carried to the midgut. The other part of the pancreas consists of structures termed the islets of Langerhans. The cells here are endocrine cells that secrete hormones into the blood. One subset of the endocrine cells, called beta (β) cells, secretes insulin. Another subset, the alpha (α) cells, secretes glucagon.
As a meal is processed, glucose, fatty acids, and other products of digestion enter the blood in abundance. Insulin is secreted at this time. It stimulates many types of cells in the body to take up glucose. In this way insulin plays two major roles. It stabilizes the blood glucose concentration, keeping it from rising too high (FIGURE 30.16). It also promotes the use and storage of glucose when glucose is available in abundance. Insulin stimulates both liver and muscle cells to synthesize glycogen from blood glucose, thereby storing the glucose for future use. Insulin also has roughly parallel effects on blood fatty acids and amino acids. It stimulates uptake of both from the blood, storage of fatty acids by formation of lipids in fat cells, and use of amino acids for protein synthesis.
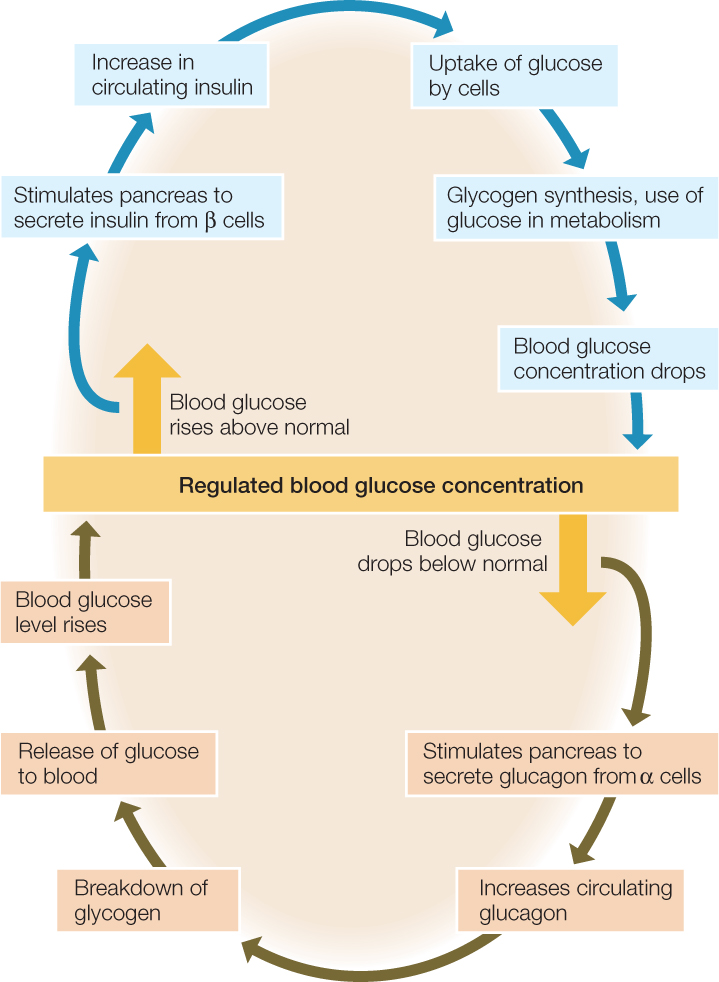
During the postabsorptive state between meals, insulin secretion stops, and the pancreas secretes glucagon. In many ways, glucagon’s effects are the opposite of insulin’s. Glucagon promotes both breakdown of glycogen into glucose molecules and release of glucose into the blood. In this way it helps stabilize the blood glucose concentration by preventing the concentration from falling too low (see Figure 30.16). It also ensures that cells throughout the body have a glucose supply even when foods are not being digested. In addition, glucagon stimulates breakdown of stored lipids and release of fatty acids into the blood.
641

Go to ANIMATED TUTORIAL 30.2 Insulin and Glucose Regulation
PoL2e.com/at30.2
CHECKpoint CONCEPT 30.5
- Blood glucose concentration is often said to be subject to negative feedback control by insulin. Explain.
- Why is it important that the endocrine cells in the gut epithelium do not all secrete the same hormone?
- Glucose is not normally found in a person’s urine. Particularly in earlier decades, glucose in the urine was used as a diagnostic indication of diabetes. In one form of diabetes, termed Type I, the ß cells in the islets of Langerhans are defective. Why would this condition lead to glucose in the urine?
Question 30.2
What are some of the specific types of organic molecules that animals must obtain in their foods, and why do they need them?
ANSWER Animals must obtain all their essential amino acids in their foods because they are not able to synthesize those amino acids and will develop deficiency diseases if they have inadequate amounts (Concept 30.1). Many types of mammals require eight amino acids, called the essential amino acids: isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan, and valine. Similarly, animals are unable to synthesize vitamins and essential fatty acids. To get adequate amounts, they must obtain these molecules in their foods. Two points should be stressed for fully understanding these concepts. First, the essential nutrients are not the same for all animals (Concept 30.1). Second, microbial symbionts sometimes provide essential nutrients, in which case animals do not literally need to eat the nutrients (Concept 30.2). Ruminants, for example, do not need to ingest certain required B vitamins—even though they are unable to synthesize them—because the microbial symbionts in their rumen synthesize the vitamins.