Concept 31.2: Animals Have Evolved Diverse Types of Breathing Organs
Some types of animals do not have specialized breathing organs. Instead, they have body plans in which most of their cells are close to a body surface. Planaria and other flatworms provide an example:

Because of a flatworm’s flat and thin body plan, O2 can diffuse directly to each cell at a rate high enough to meet the cells’ metabolic needs, simply by entering the body across an external surface. Likewise, CO2 can exit the cells and the body by diffusion.
Sponges also lack specialized breathing organs:

The bodies of sponges are organized around an extensive system of internal passageways through which water flows continually, driven by beating flagella. All of a sponge’s cells are close enough to one of these passageways to receive O2 by diffusion from the environmental water flowing through. CO2 leaves the cells in the same way.
649
Most types of animals, however, have evolved specialized breathing organs. The reason is that in most animal body plans, the great majority of cells are far from an external body surface. If we look at a snail, a lobster, a fish, or a bird, we quickly see that most cells are much farther than 0.5 millimeters from a body surface. Even if the external body surface were freely permeable to O2 and CO2, Krogh’s rule of thumb tells us that diffusion through body fluids, directly from the outside environment, could not possibly meet the needs of most cells for respiratory gas exchange.
Specialized breathing organs have large surface areas of thin membranes
In what ways are lungs and gills specialized for breathing? One of their most important features is they have large surface areas of thin gas exchange membranes. Both of these properties—large surface area and thinness—make perfect sense when we look at the diffusion equation. As you’ll recall, O2 and CO2 cross the gas exchange membrane by diffusion. The equation tells us that a large area, A, helps ensure a high rate of diffusion. The equation also tells us that a short diffusion path, L, as exists with a thin membrane, aids rapid diffusion.
Whether the breathing organs are lungs or gills, a large surface area is achieved by complex patterns of tissue folding and branching. These foldings and branchings are invaginated into the animal’s body in the case of lungs, and evaginated in the case of gills (see Figure 31.4).
Consider your own lungs. The area of the gas exchange membranes in them is about 130 square meters! That’s the same as the floor area of a classroom for 80 students. All that surface area can fit into the small volume of your lungs because of branching and folding. The branching resembles that of a tree. Thus, to understand it better, you can think of your lungs as an upside-down tree, where your trachea, or windpipe, corresponds to the tree trunk. Your trachea branches to form two tubes, then each of those tubes branches, and those branches also branch—and so on until the final branches are 23 steps removed from the trunk—the trachea. Each final branch ends in a closed sac of gas exchange membrane that is itself highly folded to increase its surface area. The result is 130 square meters of thin membrane with air on one side and blood on the other side. By virtue of both its large area and thinness, this membrane is ideally suited for O2 to diffuse rapidly into your blood and for CO2 to diffuse rapidly out.
How else are lungs and gills specialized for breathing? Another way is the presence of a ventilation mechanism. Without ventilation, our lungs would not be useful. Muscles in our thorax and abdomen expand our lungs when we inhale and sometimes help to contract our lungs when we exhale, causing a bulk flow of air in and out. This bulk flow is as essential as the gas exchange membranes for our lungs to work.
Finally, most breathing organs need circulatory specializations because cells throughout the body depend on blood circulation to receive O2 taken up in the lungs or gills. Perfusion refers to blood flow through the capillaries or other small blood vessels of a tissue. Perfusion of the gas exchange membranes in a breathing organ must be rapid enough for the blood to pick up O2 as fast as it is delivered by breathing. Then blood must circulate throughout the body at a high enough rate to deliver O2 by bulk flow from the lungs or gills to meet the O2 requirements of all the cells.
In short, most specialized breathing organs have three elements:
- A ventilation system that brings air or water rapidly to the gas exchange membrane
- A thin gas exchange membrane of large surface area, with air or water on one side and blood on the other side, for rapid diffusion
- A high rate of blood perfusion provided by the circulatory system, which also pumps blood rapidly between the lungs or gills and the rest of the body
The directions of ventilation and perfusion can greatly affect the efficiency of gas exchange
When ventilation is unidirectional, a key question is whether blood perfusion of the gas exchange membrane is in the same, or opposite, direction. Let’s consider a water-breathing animal, although the principles we’ll discuss apply whether the environmental medium is water or air. We can think of blood flow along the gas exchange membrane as being a fluid stream, and of water flow along the membrane as a second fluid stream. The entire system of blood and water flow is a cocurrent system (sometimes called a concurrent system) if the two fluid streams flow in the same direction. It is a countercurrent system if the two fluid streams flow in opposite directions. Gas exchange between blood and water is called cocurrent gas exchange in a cocurrent system and countercurrent gas exchange in a countercurrent system.
Keep in mind that as blood circulates in the body of a water-breathing animal, it flows back and forth between the gills and the systemic tissues. The systemic tissues in any animal are all the tissues other than those of the breathing organ. In the systemic tissues, the blood releases O2 to the cells and picks up CO2. Then the blood returns to the gills to unload CO2 and pick up more O2.
Also keep in mind how partial pressure affects the rate of gas diffusion. The diffusion equation shows this effect. For our present purposes, the partial pressures of interest are those in the water and blood at the gas exchange membrane. Thus we can think of P1 and P2 in the diffusion equation as being the O2 partial pressures in the water and blood. The rate of diffusion is proportional to the difference between these two partial pressures.
Now let’s follow the blood from the moment it arrives at the gas exchange membrane. At that moment, the blood has a low O2 partial pressure because it has just returned from the systemic tissues. What is the O2 partial pressure of the water the blood first encounters? In a cocurrent system (FIGURE 31.6A), this water is fresh and has a high O2 partial pressure. Thus there is a large difference in O2 partial pressure between the water and blood, and O2 diffuses rapidly into the blood. However, as the blood and water flow in the same direction along the gas exchange membrane in a cocurrent system, the O2 partial pressure of the blood rises whereas that of the water falls, so the difference in partial pressure becomes smaller and smaller. Accordingly, the rate of O2 diffusion into the blood becomes lower and lower.

650
A key question to ask is, When the blood finally leaves the gas exchange membrane, what is the O2 partial pressure of the water with which it last undergoes gas exchange? As Figure 31.6A shows, that O2 partial pressure is far lower than that in fresh environmental water. The O2 partial pressure of the blood cannot be any higher. Thus the blood O2 partial pressure is doomed to being far lower than the O2 partial pressure of fresh environmental water.
A countercurrent system works in a very different way (FIGURE 31.6B). In this system, What is the O2 partial pressure of the water that the blood first encounters? This water is stale and has a low O2 partial pressure in comparison with fresh environmental water. Thus at the spot where blood first arrives at the gas exchange membrane, there is just a moderate difference in O2 partial pressure between the water and blood in comparison to the difference in the cocurrent system, and O2 diffuses into the blood at only a moderate rate by comparison. However, this moderate difference is maintained in the countercurrent system rather than becoming smaller as it does in the cocurrent system. As the blood flows along the gas exchange membrane and its O2 partial pressure rises, the blood keeps encountering fresher and fresher water with a higher and higher O2 partial pressure. For this reason, at every step of the way the O2 partial pressure of the water is substantially higher than that of the blood. As a consequence, O2 keeps diffusing into the blood at a substantial rate, even as the blood becomes richer and richer in O2.
Now we come back to the same key question we asked earlier, When the blood finally leaves the gas exchange membrane, what is the O2 partial pressure of the water with which it last undergoes gas exchange? In the countercurrent system, this water is entirely fresh environmental water and has a high O2 partial pressure. The O2 partial pressure of the blood can accordingly rise high enough to be relatively close to that of fresh environmental water.
In conclusion, blood leaves the gas exchange membrane with a higher O2 partial pressure when exchange is countercurrent than when it is cocurrent. This difference occurs because, as blood flows through the gills, the blood takes up more O2 from the water in a countercurrent system, where uptake continues at a substantial rate every step of the way along the whole length of the gas exchange membrane. A countercurrent system is more efficient in transferring O2 from the environmental medium to the blood.
LINK
Countercurrent exchange is also important for thermoregulation in some animals; see Figure 32.16
Many aquatic animals with gills use countercurrent exchange
The gills of many aquatic invertebrates, such as clams and aquatic snails, employ countercurrent gas exchange. The gills of fish do also, and here we will look at those in detail.
The gills of a fish are supported by bone-reinforced gill arches positioned between the mouth cavity and the protective opercular flaps on the sides of the fish just behind its eyes (FIGURE 31.7A). The gill arches resemble vertical pillars with spaces—termed gill slits—in between. A fish pumps water unidirectionally into its mouth, through its mouth cavity, through its gill slits, over its gills, and out from under the opercular flaps.

Fish gills have an enormous surface area for gas exchange because they are extensively branched and folded. Each gill consists of two rows of long, flat filaments called gill filaments (FIGURE 31.7B). The upper and lower surfaces of each gill filament are folded into many flat folds—called secondary lamellae—that run perpendicular to the long axis of the filament (FIGURE 31.7C). The secondary lamellae are the principal sites of respiratory gas exchange. Blood flows through small blood passages within each lamella, and these blood passages are separated from the water flowing over the gills by just a thin epithelium. This epithelium is the gas exchange membrane. Its total surface area is enormous because of the great numbers of filaments and secondary lamellae.
651
Blood flows unidirectionally through each secondary lamella, as Figure 31.7C shows, and the direction of water flow over the lamella is opposite to the direction of blood flow. Thus efficient, countercurrent gas exchange takes place. As blood enters a lamella, it is deoxygenated, or low in O2 (see Concept 32.5). It gradually becomes oxygenated by taking up O2 by diffusion from the water. If we focus on the spot where blood leaves a lamella, we see that the water on the outside of the lamella at that spot is water that has just started to flow across the gills. That water is accordingly high in O2, ensuring that the blood is brought to the highest possible O2 partial pressure before exiting the lamella and traveling to the systemic tissues.
Most fish ventilate their gills by means of a two-pump mechanism based on contractions of breathing muscles. One pump occurs in the mouth cavity. Muscles first expand the mouth cavity so it fills with water through the mouth. Muscles then contract the mouth cavity with the mouth closed, driving the water through the gill slits. The second pump occurs in the opercular cavities—the spaces under the opercular flaps. Muscles pull the opercular flaps away from the body wall, creating a sucking force that draws water through the gill slits into the opercular cavities. Muscles then pull the opercular flaps toward the body wall, discharging water.
Some highly active fish—such as tunas and some sharks—ventilate their gills by driving water across them as they swim. A swimming tuna holds its mouth open so that water is rammed into its mouth and across its gills by the fish’s forward motion. Fish of this type can die if prevented from swimming.
Most terrestrial vertebrates have tidally ventilated lungs
The lungs of terrestrial vertebrates evolved as saclike outpocketings of the digestive tract. This ancient heritage is still reflected today in the fact that, except in birds and some crocodilians, the finest branches of the tubular parts of the lungs end in closed, dead-end sacs of gas exchange membrane. Because of this structure, ventilation of the gas exchange membranes cannot be unidirectional but must be tidal: fresh air flows in and exhaled gas flows out by the same route. Without unidirectional ventilation, cocurrent and countercurrent gas exchange are not possible.
652
As we already noted, there are 23 levels of branching in the lungs of humans. Other mammals have similar levels of complexity. Amphibian lungs are much simpler and help us understand what vertebrate lungs probably were like early in their evolution. An amphibian lung has an open central cavity, which is surrounded by intricately folded, well-perfused membranes:
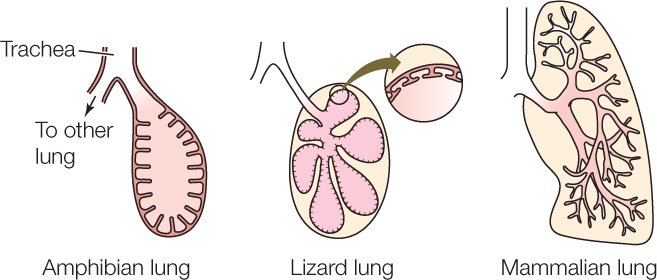
Compared with amphibians, some lizards exemplify increasing complexity in that each lung is subdivided into many separate parts served by separate airway tubes, instead of having a single, open central cavity (see above). The intricate branching of airways in the mammalian lung represents a further, more elaborate step in the same direction. The area of gas exchange membranes in the lungs of a mouse or rat is about ten times greater than the area in a mouse-size or rat-size lizard.
The inside surfaces of vertebrate lungs are coated with a thin layer of metabolically produced lipids and proteins called lung surfactant. By reducing surface tension at the surfaces of the lung epithelium, this material helps stabilize the complex three-dimensional structure of the gas exchange membranes. Infants born without adequate lung surfactant (a problem most often seen in premature infants) are in danger of dying unless they receive medical care because the interior surfaces of their lung membranes tend to stick together, requiring lots of effort to pull them apart with each inhalation.
In amphibians, the lungs are inflated by positive pressure. During each inhalation, the mouth cavity is first filled with air from the atmosphere. The mouth and nostrils are then sealed, and muscles contract the mouth cavity, raising the pressure high enough to drive air into the lungs. This mechanism is the same as that used by air-breathing fish (such as lungfish) to fill their air-breathing organs, suggesting that amphibians carry over the mechanism from their fishlike ancestors.
With the evolution of reptiles, a major transition in ventilation took place. Air is drawn into the breathing system by suction. This is true in today’s reptiles, including birds. It is also true in mammals. In most non-avian (non-bird) reptiles and in mammals, the lungs are expanded during inhalation, creating a lower air pressure inside the lungs than in the atmosphere, thereby sucking air into the lungs (see Concept 31.3 for greater detail in humans).
Birds have rigid lungs ventilated unidirectionally by air sacs
Birds and mammals both have high metabolic rates and thus require sophisticated lungs that can supply O2 at high rates to the rest of the body. The lungs of birds and mammals, moreover, are both believed to have evolved from lungs similar to those seen in today’s non-avian reptiles. The lungs of birds, however, are radically different from those of mammals.
The lungs of birds are often described as “rigid.” This does not mean they are solid or hard, but rather that they do not change much in volume as a bird inhales and exhales.
The breathing system of a bird includes air sacs as well as lungs (FIGURE 31.8A); the air sacs are the parts that undergo large changes in volume as a bird inhales and exhales. These thin-walled sacs connect to airways in the lungs and fill much of the internal space in a bird’s body. The air sacs do not participate in exchange of O2 and CO2 between the air and blood. Instead, their function is ventilation. They expand and contract during inhalation and exhalation, acting like bellows and forcing air to flow past the gas exchange membranes in the rigid lungs.
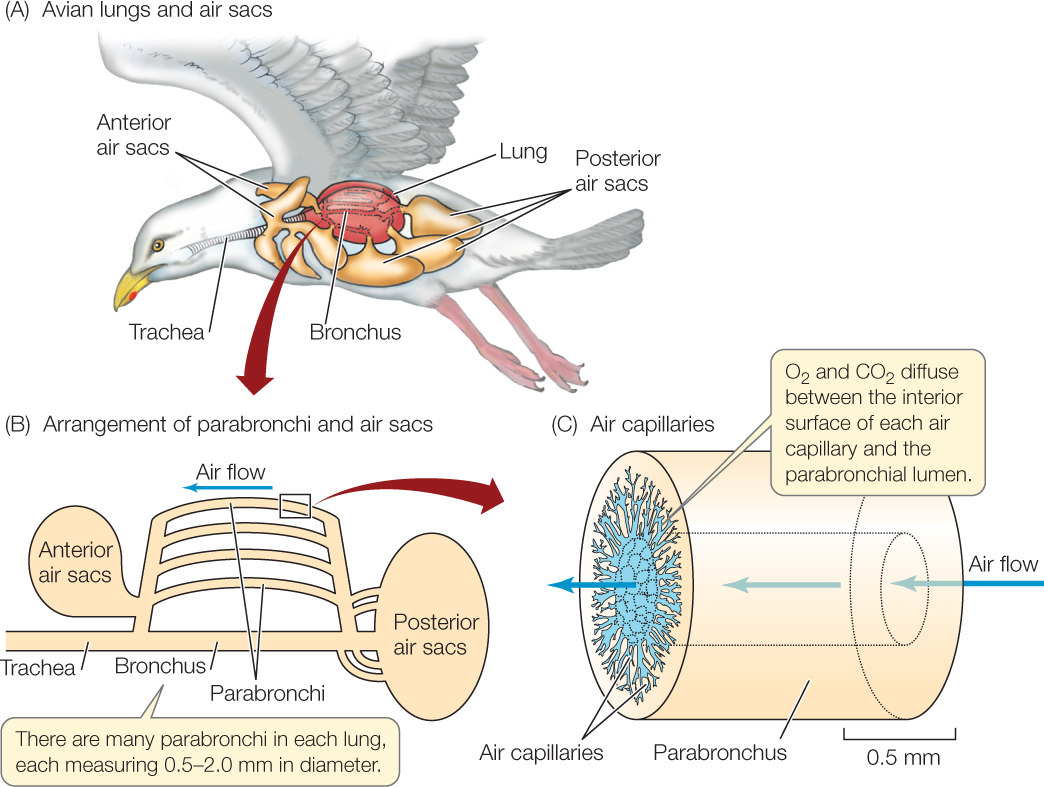
653
Long, tiny tubes called parabronchi (singular parabronchus) are the most important airways for gas exchange in the lungs of a bird (FIGURE 31.8B). As astounding as it may sound, air flows through most of a bird’s parabronchi unidirectionally, from posterior to anterior, during all phases of its breathing cycle, whether the bird is inhaling or exhaling! Air flows unidirectionally through the parabronchi because of coordinated actions of thoracic and abdominal muscles that expand and contract the anterior and posterior sets of air sacs (FIGURE 31.9). When a bird inhales, some of the incoming fresh air travels to the posterior of each lung and enters the parabronchi. That air then flows through the parabronchi from posterior to anterior and exits into anterior air sacs. Some of the incoming fresh air also inflates the posterior air sacs. Then, when the bird exhales, this fresh air in the posterior air sacs is driven into the parabronchi, again flowing from posterior to anterior. As this occurs, stale air exiting the parabronchi at the anterior end, plus stale air from the anterior air sacs, is exhaled into the atmosphere.
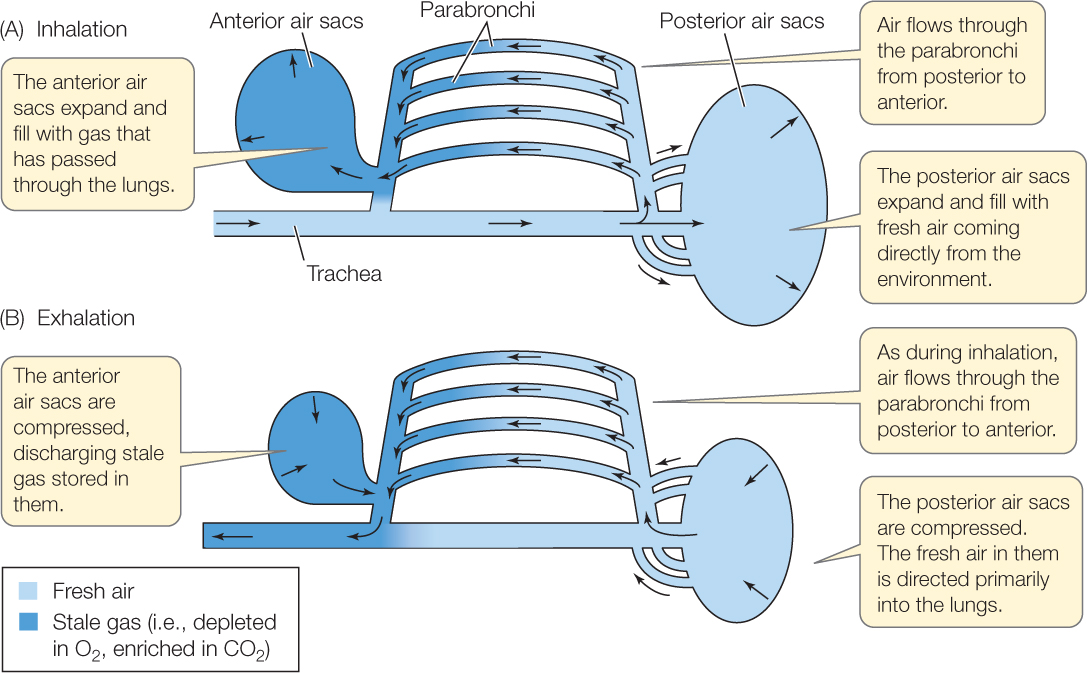

Go to ANIMATED TUTORIAL 31.1 Airflow in Birds
PoL2e.com/at31.1
Tiny air capillaries extend into the walls of each parabronchus from the parabronchial lumen (FIGURE 31.8C). These air capillaries are so minute that if the openings of 1,000 of them were lined up in a row, they would cover only 1 centimeter or less. The walls of the air capillaries are richly perfused with blood and serve as the gas exchange membranes. As air moves by bulk flow through the parabronchi, O2 and CO2 are thought to move in and out of the tiny air capillaries by diffusion. Gas exchange with the blood is termed “cross-current”—a type of exchange intermediate in efficiency between cocurrent and countercurrent.
Insects have airways throughout their bodies
In insects (and spiders), a breathing system has evolved that is unlike that of any other animals (FIGURE 31.10). This breathing system is a type of lung because it consists of gas-filled tubules that are invaginated into the body from the body surface. The tubules branch everywhere in an insect’s body. In fact they branch in such fantastically intricate ways that a gas-filled tubule comes close to every cell. In some cases tubules grow into cells, indenting the cell membrane inward. The tubules connect to the atmosphere at several openings, called spiracles, on the outside surface of an insect’s body. O2 makes its way directly from the atmosphere, through the gas-filled tubules, to each cell. CO2 moves directly from all cells to the atmosphere. Because of this unique breathing system, blood flow is generally unimportant in delivering O2 and getting rid of CO2.

Most of the gas-filled tubules are called tracheae, and for this reason, insects are said to have a tracheal breathing system. The tracheae are lined with a thin, gas-permeable layer of chitin, which provides support and keeps them from collapsing shut. The finest tubules are called tracheoles. Air sacs are connected to the tracheae in many insects.
The air inside the tracheal breathing system is believed to be completely still in some insects. In these cases O2 and CO2 travel everywhere by diffusion. Keep in mind that Krogh’s rule of thumb refers just to diffusion through water. Diffusion in air is much faster (200,000 times faster in the case of O2) because random molecular movements are more vigorous in gases than in water. Because of the fast diffusion that can occur in air, insects can be moderately large (e.g., 0.5 cm long) and still have their gas exchange needs met by diffusion in the tracheal system, provided they are not very active.
In insects that are large or active, mechanisms have evolved to create bulk flow of air inside the tracheal system. Grasshoppers, for example, pump air through their tracheae by contracting and elongating their bodies. Bees use air sacs as bellows to drive air through their tracheae. The tracheae throughout an insect’s body sometimes undergo steady, rhythmic contractions.
654
CHECKpoint CONCEPT 31.2
- What process is responsible for the movement of O2 into a flatworm’s body?
- As blood flows along the gas exchange membrane, how does the difference in O2 partial pressure between the environmental medium and blood vary in countercurrent and cocurrent systems?
- Why can’t diffusion alone supply fresh O2 molecules to the breathing systems of many organisms?
- Mammals, birds, and insects are the three groups of animals in which species with the highest known metabolic rates occur. Of these three groups, two—mammals and birds—have high-performance circulatory systems that can circulate blood rapidly throughout the body and perfuse tissues at high rates. Insects do not have high-performance circulatory systems. Explain.
Now let’s take a closer look at the anatomy and function of mammalian lungs, focusing on the human breathing system.