Concept 32.4: Many Key Processes Occur in the Vascular System
All the blood vessels, considered together, make up the vascular system. The circulatory system is sometimes called the cardiovascular system to emphasize that it consists of the heart plus the blood vessels.
The vascular system is an elaborate system in which the vessels in various parts are specialized to perform specific functions. Let’s briefly review the vessels in the human circulatory system in the order that blood flows through them.
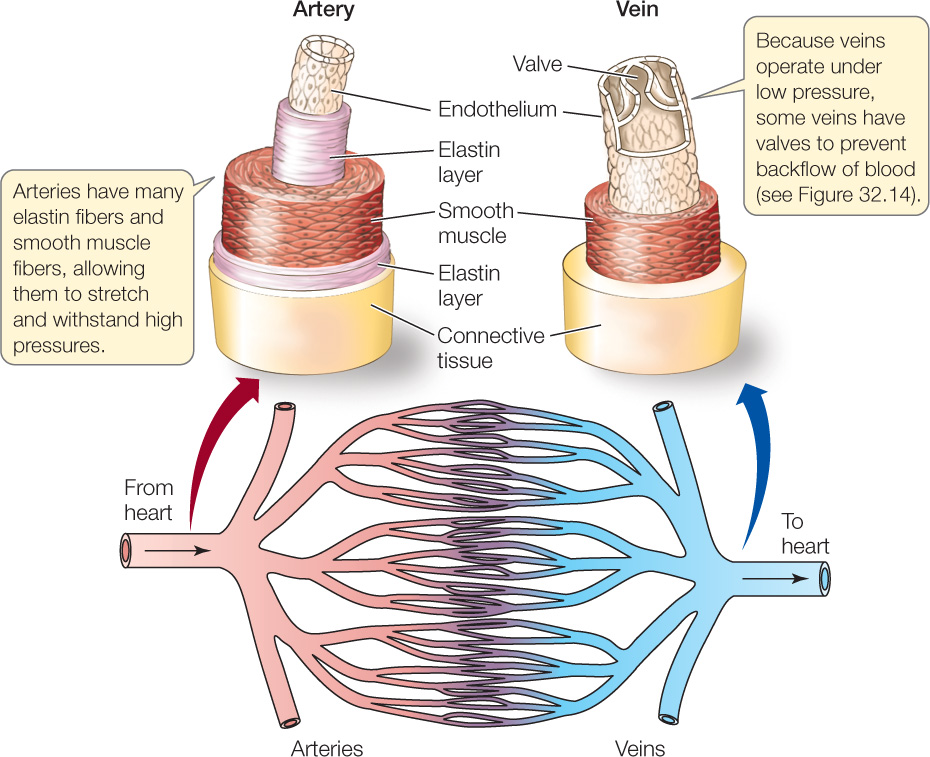
The arteries must withstand high pressures. To do this, they have thick walls composed of elastic fibers and layers of smooth muscle (FIGURE 32.12). The elasticity of the arteries allows them to play two critical, interrelated roles. Each time the heart beats, the arteries stretch a bit, dampening the pressure surge and storing in their stretched state some of the energy imparted to the blood by the heart. Then, during the next diastole, the stretched arteries return to their unstretched state, in the process squeezing the blood and helping maintain blood pressure while the heart is between beats. In this way the arteries play a pressure-dampening role and a pressure-storage role. Were it not for these processes, arterial blood pressure would vary much more than it does between systole and diastole, shooting sharply up and plummeting sharply down. The arteries branch and rebranch, carrying blood to all organs and tissues.
Go to ACTIVITY 32.3 Structure of a Blood Vessel
PoL2e.com/ac32.3
The blood then flows into microcirculatory beds where the arterioles and capillaries play the vital roles we discussed in Concept 32.1. Tiny arteries and the arterioles exercise vasomotor control over the flow of blood to the capillary beds. The capillaries are the principal sites of exchange of materials between the blood and the cells of the tissues.
Blood then enters tiny veins, which merge progressively with other veins, finally leading to the great veins that enter the heart. Blood pressure is far lower in the veins than in the arteries, and the walls of the veins are thinner and expand more easily (see Figure 32.12). A major role of the veins is to serve as a blood volume reservoir. Any blood that is not held elsewhere in the circulatory system is held in the veins.
Pressure and linear velocity vary greatly as blood flows through the vascular system
The principal driving force for blood to flow through the vascular system is the pressure imparted to the blood by the beating heart. The blood pressure represents potential energy. This potential energy is converted to energy of motion as the blood flows, and then it is lost as heat through a type of friction that occurs during blood flow. As a result, the blood pressure falls as the blood flows. Moreover, the pressure falls in direct proportion to the amount of work that must be done to make the blood flow.
Fluids flow easily through large-diameter tubes. For this reason, not much work needs to be done to cause blood to flow through the large, or even medium-sized, arteries. With little work needing to be done, the blood pressure falls only moderately as blood flows through the beginning branches of the arterial vasculature (FIGURE 32.13).
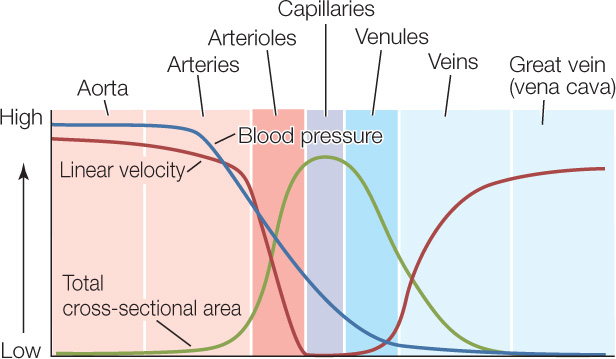
In dramatic contrast, a great deal of work needs to be done to make blood flow through the microscopically tiny vessels of the microcirculation. This work results in a precipitous drop in blood pressure as blood flows through the arterioles, capillaries, and venules (see Figure 32.13). The loss of pressure is so great that, by the time the blood flows into the veins, its pressure is only a little higher than zero.
674
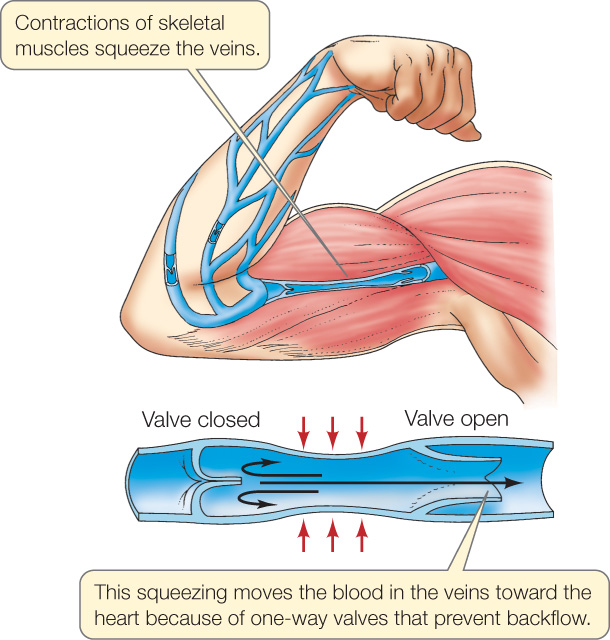
The low blood pressures in the veins help explain why one-way valves have evolved in the veins (valves do not occur in arteries). The pressure from the beating of the heart is reduced to such a low level in the veins that it may not always be sufficient to make blood flow back to the heart. With one-way valves, contractions of skeletal muscles can help propel blood to the heart (FIGURE 32.14).
Like blood pressure, the linear velocity of the blood—which is simply the linear distance the blood travels per second—also changes dramatically as blood travels through the vascular system (see Figure 32.13). Suppose that a river suddenly divides into 10,000 little streams that collectively have a cross-sectional area much greater than the cross-sectional area of the river. We all appreciate that water will flow more slowly after it enters the little streams. This same logic applies in the circulatory system. Individual capillaries have minute cross-sectional areas. The capillaries are so numerous, however, that their collective cross-sectional area is hundreds of times greater than the cross-sectional area of the aorta (see Figure 32.13). Accordingly, as blood flows from the aorta to the capillaries, it slows from an astounding linear velocity of more than 40 centimeters per second to almost zero.
Why does this slowing matter? Capillaries are less than 1 millimeter long. For the blood to exchange materials with the tissues, it must be moving very slowly as it passes through them.
Animals have evolved arrangements of blood vessels that help them conserve heat
When arctic foxes (FIGURE 32.15) walk over the snow and ice in winter—or when arctic gulls stand on winter ice—the tissues in their foot pads and foot membranes are at temperatures near 0°C, even though the tissues in the core of their body (the thorax and abdomen) are at almost 40°C. By having near-freezing tissue temperatures in their feet, the animals conserve energy because they do not lose much heat across their feet. They also keep their feet dry because the tissues of their feet are too cold to melt the snow or ice. They do not risk getting frostbite because the temperature of their feet is highly controlled.

Heat is easily lost across body appendages of all kinds, including not only the legs and feet of mammals and birds, but also the tails and ear flaps (pinnae) of mammals. Many species have evolved ways to let the tissues in the outer ends of these appendages cool off as a way of conserving heat in their body core in cold environments.
How is the cooling of the tissues achieved? The cells in the appendages need O2, so blood flow to the appendages can’t simply be stopped to keep body heat from entering them.
Many types of mammals and birds have evolved arrangements of arteries and veins that allow them to conserve heat by countercurrent heat exchange. As we discuss this process, keep in mind that the blood vessels involved are arteries and veins—not capillaries. The walls of arteries and veins are far too thick for O2 or other dissolved materials to pass through them. Heat, however, can pass through.
Figure 32.16A shows two arrangements of arteries and veins in an appendage. The diagrams show only one artery and one vein, but that’s just for simplicity. In the arrangement at the left, the arteries and veins are far away from each other and do not interact. At the right, the arteries and veins are touching. The right-hand arrangement is termed countercurrent—a term that refers to two closely juxtaposed fluid streams flowing in opposite directions. In the countercurrent arrangement, heat can move from one fluid stream to the other. Countercurrent heat exchange can therefore occur.
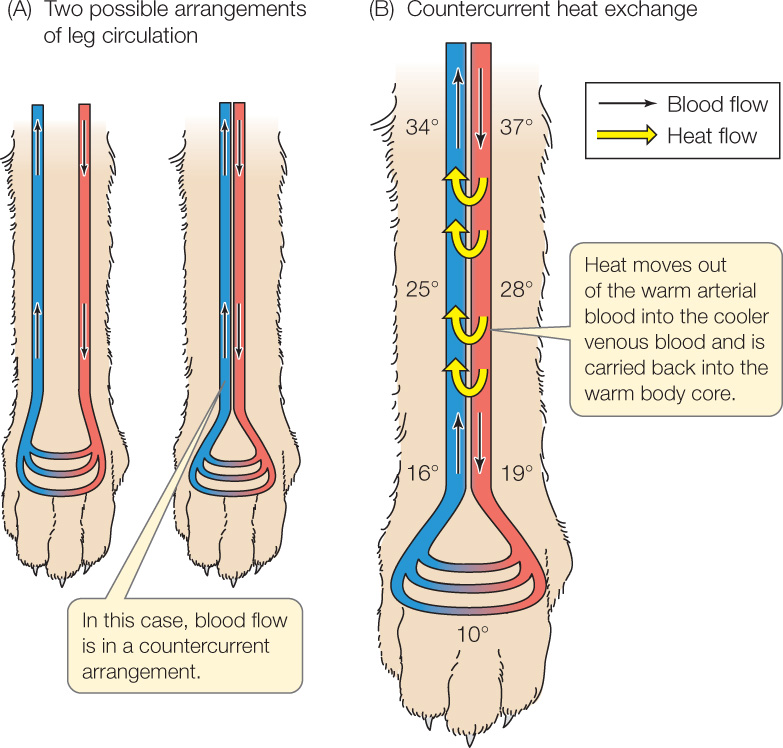
What happens in an appendage with countercurrent heat exchange? As warm, arterial blood flows into the appendage, the warm blood comes close to venous blood that is leaving the appendage. This venous blood is cooler because it is returning from the far end of the appendage. Heat travels out of the warm arterial blood into the cooler venous blood (Figure 32.16B). This process continues as the arterial blood flows farther into the appendage—because even though the arterial blood gets cooler, the venous blood next to it also gets cooler. In this way, a lot of heat moves out of the arterial blood and into the venous blood in the upper parts of the appendage. Moreover—and this is very important—this heat is immediately transported back into the animal’s body core as the venous blood flows back into the body. This heat is never carried by the blood to the outer end of the appendage, and therefore cannot be easily lost to the cold environment. The outer end of the appendage remains at a low temperature because the heat never gets to it.
675
The effectiveness of countercurrent heat exchange depends on how extensively the arteries and veins make contact with each other. With extensive, highly controlled interaction between the two sets of vessels, the tissues at the outer end of an appendage can be kept just above the freezing point, as in arctic foxes.
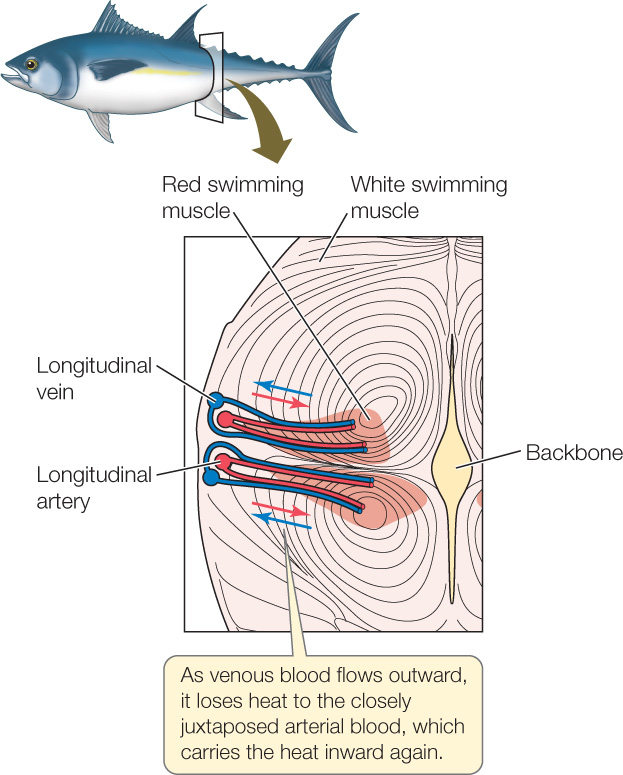
Tuna also use the countercurrent principle to keep their red swimming muscles warm (FIGURE 32.17). The red muscles are positioned in a very unusual place in tuna, deep in their body, nearer their backbone than their skin (see Figure 33.14). Tuna also have evolved a very unusual arrangement of arteries and veins. The longitudinal arteries and veins that carry blood along the length of their body are just under their skin. Branches from each longitudinal artery travel from the skin inward to the red swimming muscles. Veins from the red muscles travel outward to connect with a longitudinal vein. These arteries and veins—which are carrying blood to and from the red muscles—are closely juxtaposed and exceedingly numerous. Because the red swimming muscles work hard, they produce heat at a relatively high rate. The venous blood, flowing outward from the muscles, tends to carry this heat away. However, the arteries carrying inward-flowing arterial blood are touching the veins. Heat from the venous blood thus moves into the arterial blood and is carried directly back to the swimming muscles. Because of this countercurrent heat exchange, the muscles retain the heat they make and are warmed by it.
Blood flow leaves behind fluid that the lymph system picks up
Most blood capillaries are so permeable that at their high-pressure arterial end, blood pressure squeezes fluid out of the blood in the capillaries and into the surrounding interstitial fluids. Before the blood leaves capillaries, it takes up some of this fluid again, but not all of it. These processes were first clarified by the physiologists E. H. Starling and E. M. Landis.
Starling and Landis realized that blood in the capillaries has a higher osmotic pressure (because it has more dissolved protein) than the interstitial fluids (see Concept 5.2). For this reason, water tends to be drawn into the blood in a capillary by osmosis across the capillary walls. Water also tends to be driven out of the blood in a capillary by the high blood pressure inside the capillary. At the beginning of a capillary, where blood has just entered it, the effect of the blood pressure exceeds the effect of the osmotic pressure, and fluid moves out of the blood (FIGURE 32.18). Blood pressure falls, however, as blood flows through a capillary. At the end of the capillary, therefore, the effect of the osmotic pressure exceeds the effect of the blood pressure, and fluid moves from the interstitial fluids into the blood.
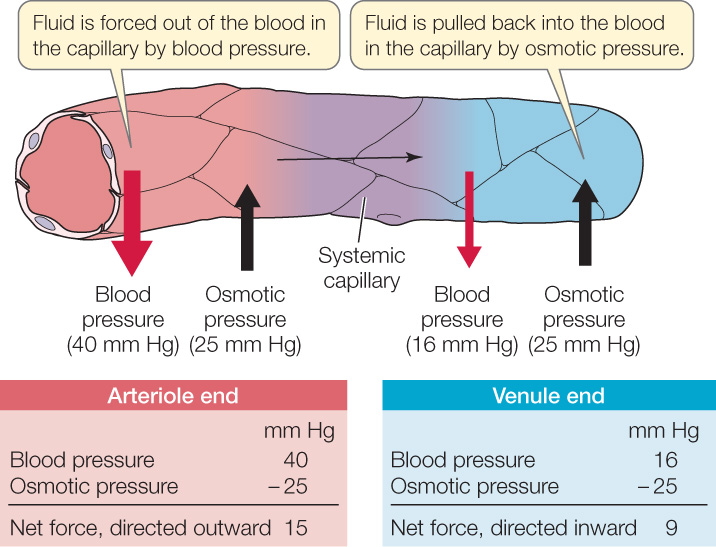
676
The net effect of these dynamic processes is a small loss of fluid from the blood. What is the fluid loss per day in adult humans, considering all the capillaries in the body and all the times that blood passes through them? Blood flow through the systemic capillaries leaves behind about 2–4 liters of fluid in a person’s systemic interstitial fluids each day.
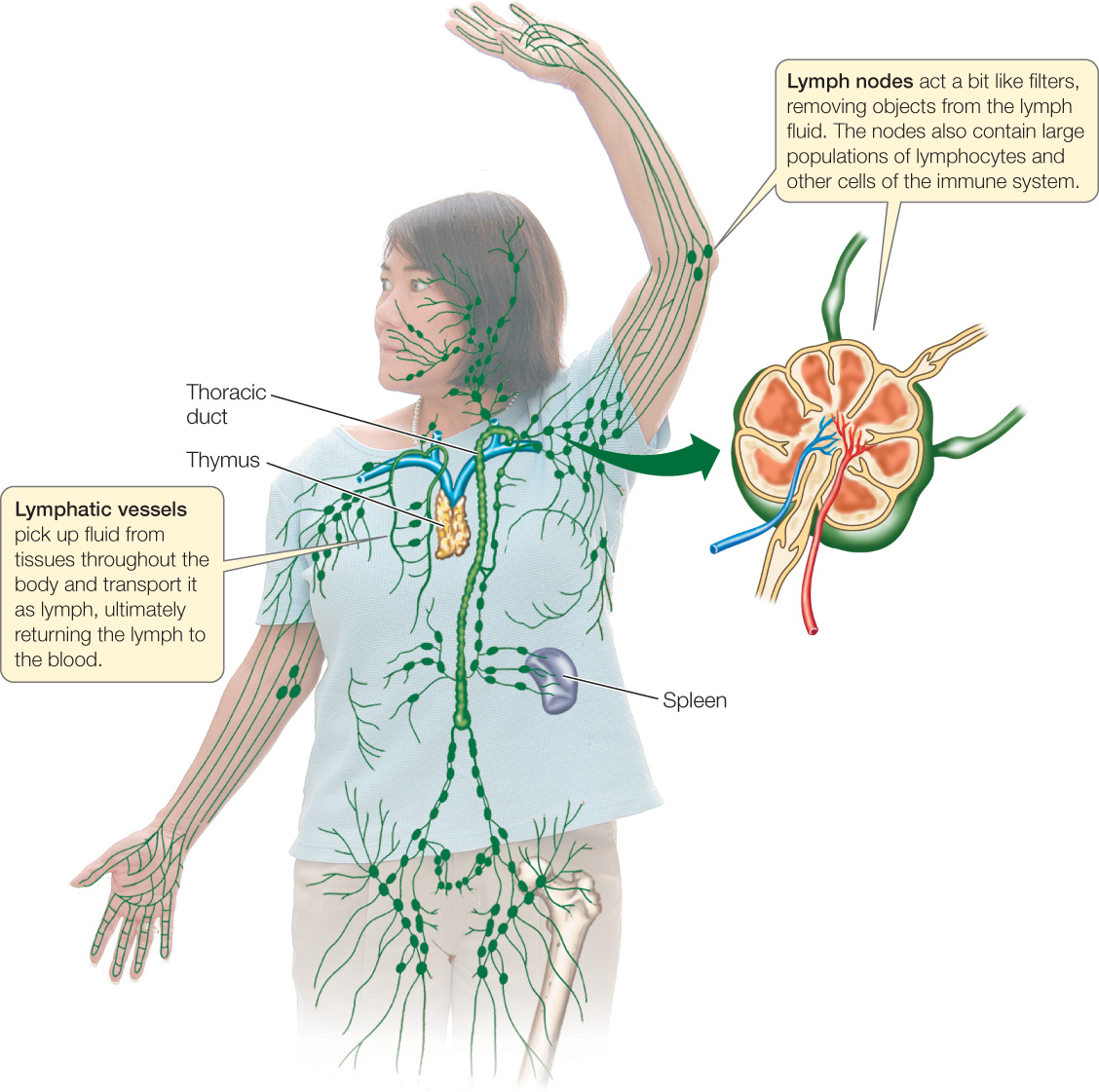
The lymphatic system collects this excess fluid (called lymph) and returns it to the blood. The lymphatic system is an elaborate system of vessels that is separate from the blood vascular system (FIGURE 32.19). Tiny lymphatic capillaries found in tissues throughout the body merge into progressively larger lymphatic vessels. Finally, certain of the lymphatic vessels connect with veins at the base of the neck, returning collected fluid to the blood.
CHECKpoint CONCEPT 32.4
- In which vessels of the circulatory system does the blood move most slowly? Why, and why is this important?
- Certain parasitic worms can block lymphatic vessels. Suppose the lymphatic vessels draining a person’s leg were completely blocked by a parasite infestation. What do you think would happen to the leg? Explain.
- Mammals and birds have very high aortic blood pressures in comparison with other animals, and the heart must work hard to maintain these pressures. Why must the blood pressures be so high?
677
Let’s turn now to the fluid that the heart is pumping: the blood. What are the major components of blood, and what are their major functions?