Concept 33.3: Skeletal Muscle Performance Depends on ATP Supply, Cell Type, and Training
Skeletal muscles vary in performance from one muscle to another and from time to time. Our postural and finger muscles illustrate differences among muscles. Our postural muscles sustain loads steadily over long periods of time, whereas our finger muscles typically contract quickly for brief periods.
Muscle power output depends on a muscle’s current rate of ATP supply
Muscles perform work, by which we mean that they apply forces to bones or other structures—forces that often make the structures move. For understanding the performance of work by a muscle cell, the cell’s fuel supply for work is often as important as the properties of its contractile apparatus. Muscle cells depend on ATP as their source of energy to do work. For this reason, their rate of doing work—their power output—at a given time can be only as high as the rate at which they supply their contractile apparatus with ATP.
Muscle cells have three systems for supplying the ATP they need for contraction. Here we term these the immediate, glycolytic, and oxidative systems:
- The immediate system uses preformed ATP. This ATP is already present in a cell when contraction begins. Some preformed ATP is present literally as ATP, but muscle cells can also store small amounts of preformed ATP in chemically combined form, such as in the compound creatine phosphate found in vertebrate skeletal muscles.
- The glycolytic system synthesizes ATP during the process of contraction by anaerobically breaking down carbohydrates (see Concepts 6.3 and 29.4). In vertebrates, crustaceans, and some other animals, this system produces lactic acid. It does not require O2.
- The oxidative system also synthesizes ATP during the process of contraction, but unlike the glycolytic system, it employs aerobic breakdown of food molecules to synthesize ATP (see Concept 6.2). It requires O2 because it depends on the citric acid cycle and electron transport, fully oxidizing carbohydrates and lipids to H2O and CO2.
These three systems vary greatly in how rapidly they can produce ATP and in how long they can continue to make ATP at a high rate. Accordingly, the power output and endurance of a working muscle cell vary with which system the cell is using to make ATP (FIGURE 33.13).

When a muscle cell starts to do work, the immediate system can supply ATP extremely rapidly, enabling the cell to achieve a very high power output. The immediate system can do this because it uses ATP made at an earlier time and already present in a cell when work starts. Only a small amount of preformed ATP can be present in a cell, however. Thus the immediate system can function for only a brief period of time.
The glycolytic system can synthesize ATP at a very high rate, although not as high a rate as that at which the immediate system can supply ATP. However, the glycolytic system is self-limiting. For poorly understood reasons that may be related to the lactic acid it produces, the glycolytic system cannot keep making ATP indefinitely. As the glycolytic system makes ATP, it reaches a point at which it can no longer function, and then it must rest for a long time (tens of minutes or hours) before it can be used again to supply ATP.
692
The oxidative system makes ATP at a slower rate than the other two systems. However, its products—besides ATP—are simply H2O and CO2, which healthy animals can deal with effectively. Thus the oxidative system is not self-limiting and can continue making ATP for very long periods of time if a cell’s supply of O2 is sufficient for it to function.
A muscle cell that is capable of using all three systems of ATP supply varies in its performance, depending on which system it is using. To explore this important point, let’s return to Figure 33.13. Keeping in mind that the power output of a contracting muscle cell depends directly on its rate of ATP supply, we see that if we focus on power output, the three systems of ATP supply rank as follows: the immediate system permits a muscle cell to reach highest power output, the glycolytic system ranks next, and the oxidative system permits the lowest power output. However, if we focus on endurance, the three systems rank in the opposite order: the immediate system permits the lowest endurance, the glycolytic system ranks next, and the oxidative system permits the greatest endurance.
These principles explain why, in human athletic competition, speed and endurance are inversely related. Sprints, such as the 100-yard dash, are run at a very high speed. They depend greatly on the immediate system for ATP, which permits very high power output. The immediate system cannot supply ATP for very long, however, meaning sprints must be of short duration. Events like the half-mile run and mile run are performed at lower speeds and last longer than sprints. A key reason for these contrasts is that the half-mile and mile runs require too much ATP, in total, to be run with just the immediate system as their source of ATP. Thus they depend to a great extent on the glycolytic system, which permits muscle cells to have greater endurance than the immediate system but restricts performance to a lower power output and speed. A marathon requires far too much total ATP to be run with just the immediate and glycolytic systems as ATP sources. Only the oxidative system—based on O2 use—can provide the amount of ATP a marathon requires. The speed of running in a marathon must therefore be lower than in races fueled by the immediate and glycolytic systems, although long endurance is possible.
Long-endurance casual sports such as jogging and swimming laps depend almost entirely on the oxidative system. This is true because the oxidative system can accelerate its ATP production quickly at the start, and then can meet all the ATP needs of these forms of exercise on a minute-by-minute basis, indefinitely. This is why these sports are described as aerobic.
Muscle cell types affect power output and endurance
All muscle cells are not equal in their capability to use the glycolytic and oxidative systems to synthesize ATP. Some cells principally use the glycolytic system, and others principally use the oxidative system. There are no cells, however, that depend principally on the immediate system, which can supply only small amounts of ATP.
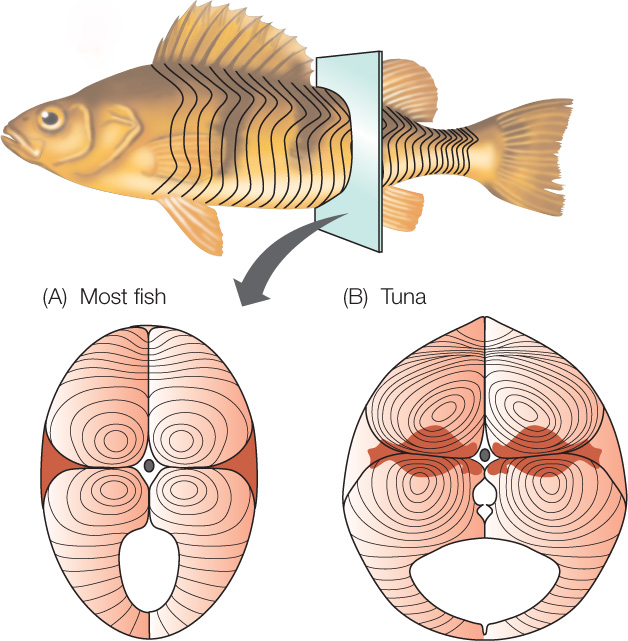
Multiple muscle cell types typically occur within a single species of animal. Two major types are often recognized: slow oxidative cells (sometimes called slow-twitch cells) and fast glycolytic cells (fast-twitch cells). In vertebrates, these cell types are also called “red” and “white.” Cells of the two types typically occur mixed together in the skeletal muscles of mammals. In some other animals, however, certain skeletal muscles are composed almost entirely of one muscle cell type or the other. For example, many fish have distinct masses of muscle that are visibly red or white (FIGURE 33.14). The red muscles are composed mostly of slow oxidative cells. The white muscles consist mostly of fast glycolytic cells.
Slow oxidative and fast glycolytic muscle cells differ in three main ways:
- Their principal mechanisms for making ATP
- Their mechanisms for taking up O2 from the blood
- Their molecular forms of myosin; the two types of cells express different genes for myosin, and their molecular forms of myosin differ in ATPase (ATP-splitting) activity
Slow oxidative cells are poised to make most of their ATP aerobically using the oxidative system of ATP synthesis. These cells need lots of O2. This explains why, in vertebrates, these cells have evolved high levels of the red, hemoglobin-like compound myoglobin in their sarcoplasm (making them red). Myoglobin speeds entry of O2 into cells from the cell surfaces. Slow oxidative cells also have large numbers of mitochondria and are rich in mitochondrial enzymes, notably the enzymes of the citric acid cycle and electron transport system (see Concept 6.2). They are not well endowed with the enzymes required for the glycolytic system to make ATP anerobically, however. Slow oxidative cells thus make most of their ATP aerobically. Accordingly, they can sustain ATP production for long, continuous periods of time, but their peak rates of ATP production are only moderately high (see Figure 33.13). Slow oxidative muscle cells also have molecular forms of myosin with relatively low ATPase activity, meaning they recycle actin–myosin cross-bridges at relatively low rates.
693
Because of their limited rates of ATP production and myosin properties, slow oxidative cells tend to contract and develop tension slowly (accounting for the word “slow” in their name). They are well suited for maintaining relatively low power output for sustained periods.
Fast glycolytic cells contrast with slow oxidative cells in all the ways we’ve mentioned. They have high levels of the enzymes needed for glycolytic, anaerobic ATP synthesis and thus can produce ATP very rapidly. They also have molecular forms of myosin with relatively high ATPase activity, meaning they can recycle actin–myosin cross-bridges rapidly. Because of their high rates of ATP production and myosin properties, fast glycolytic cells tend to contract and develop tension rapidly (thus they are “fast”). They also fatigue rapidly, however. They have relatively few mitochondria, and they lack myoglobin, so they are not red.
Fish use principally their red muscles, composed mainly of slow oxidative cells, during routine cruising, when they move slowly for long periods. They use principally their white muscles, composed mainly of fast glycolytic cells, during burst swimming, when they swim at high speeds to evade danger or catch prey. Their white muscles enable them to achieve very high power outputs for brief periods.
Individual people vary greatly in the proportions of red and white cells in their major muscles. These differences generally do not matter in sports done for fun. At the level of national or international competition, however, researchers know from many studies that champion long-distance runners, swimmers, and cyclists—who require endurance—tend to differ from champion weight-lifters, wrestlers, and sprinters—who require high power outputs for brief periods. The endurance athletes tend to have leg and arm muscles consisting mostly of slow oxidative muscle cells (FIGURE 33.15). The other athletes tend to have muscles with a high percent of fast glycolytic cells.

Training modifies muscle performance
Skeletal muscle has a remarkable ability to change its properties with use. In this way, muscle provides dramatic examples of phenotypic plasticity (see Concept 29.5). When a muscle changes in response to use, the change is often termed a “training effect.” We observe a familiar training effect when we lift weights or do other similar forms of exercise. Our muscles gradually become visibly larger. The number of muscle cells in a muscle does not change under these conditions. Instead, a muscle becomes larger because its already-existing cells increase in size (termed hypertrophy) by increasing their content of actin and myosin.
694
To fully understand training effects, two types of exercise need to be distinguished. Endurance exercise (endurance training) refers to steady, long-duration exercise such as cross-country running or long-distance cycling. Resistance exercise (resistance training) refers to exercise that generates particularly large forces, typically in relatively few repetitions spread over a short period of time. Weight-lifting and wrestling are forms of resistance exercise.
When people or research animals engage in training programs, endurance exercise causes their muscle cells to increase their numbers of mitochondria. It also causes some cells to transform from being fast glycolytic to being slow oxidative cells. It stimulates growth of additional blood capillaries in muscles—an effect that reduces the average distance between each muscle cell and O2-carrying blood in its nearest capillary. All of these training effects enhance capabilities for steady, sustained forms of exercise. Recent studies have shown that the expression of more than 100 genes is dramatically increased in muscle cells during endurance training in people or laboratory animals. Conversely, more than 100 genes are silenced when a person’s or animal’s life is entirely sedentary. Changes in gene expression help control the changes in muscle properties that occur (hormonal and metabolic control mechanisms also play roles). Investigators have been searching for transcription factors that are upregulated by endurance exercise and that may help regulate expression of genes that control transformation of cells from fast glycolytic to slow oxidative (FIGURE 33.16).
Investigation
HYPOTHESIS
Activation of PPARδ is sufficient to drive formation of slow oxidative muscle fibers.
METHOD
- Engineer a gene that encodes a permanently activated form of PPARδ.
- Produce transgenic mice that synthesize the permanently activated form of PPARδ in skeletal muscle.
- Test the transgenic mice for (a) muscle characteristics and (b) endurance. Error bars represent 1 standard error of the mean; the t-test was used to produce P-values (see Appendix B).
RESULTS

CONCLUSION
Activated PPARδ induces formation of slow oxidative muscle fibers and enhances endurance.
ANALYZE THE DATA

In a related experiment, using different sets of individual mice, the researchers tested the endurance of mutant mice that had no PPARδ transcription factor (see graph).
- Based on these data, what can you conclude about the effect of PPARδ on fatigue resistance of muscle?
- What would you expect to find if you examined cross sections of skeletal muscle tissue from the mutant mice?
Go to LaunchPad for discussion and relevant links for all INVESTIGATION figures.
aY.-X. Wang et al. 2004. PLoS Biology 2(10): e294.
When people or research animals engage in training with resistance exercise, as we have already seen, the training causes their muscle cells to build a bigger contractile apparatus, enabling the cells to better develop the forces required for weight-lifting or other resistance activities. Resistance exercise also causes some muscle cells to transform from slow oxidative to fast glycolytic.
695
CHECKpoint CONCEPT 33.3
- What factors prevent muscle cells from continuously using ATP provided by the immediate system to power contractions?
- Consider a muscle cell that contains myosin proteins specialized for rapid recycling of actin–myosin cross-bridges. Is the cell likely to make most of its ATP by using the glycolytic system or the oxidative system?
- Weekly measurements of an athlete’s gastrocnemius (calf) muscle reveal that the density of capillaries in the muscle and the number of mitochondria per muscle cell are increasing. Would you predict that the athlete has been training for a lengthy marathon or lifting weights at the gym?
Skeletal muscles, as we have seen, are enormously important. We breathe using skeletal muscles, and the skeletal muscles are responsible for most behaviors, including athletic performance. Skeletal muscle is not the only type of muscle, however, and next we will look at the other types.