Concept 37.2: The Mammalian Reproductive System Is Hormonally Controlled
Hormones play critical roles in almost every aspect of mammalian sexual reproduction. Follicle-stimulating hormone (FSH) and luteinizing hormone (LH), secreted by the anterior pituitary gland, are key players in both sexes (see Concepts 35.3 and 35.4). They are named for their roles in females, but the same hormones are secreted in males and exert major effects on tissues of the male reproductive system. Secretion of FSH and LH is controlled by neurosecretory cells in the hypothalamus of the brain. These cells secrete gonadotropin-releasing hormone (GnRH) into blood that travels to the anterior pituitary via the hypothalamo–hypophysial portal system (see Figure 35.7). In the pituitary, GnRH regulates the cells that secrete FSH and LH.
Somatic cells in the gonads produce steroid sex hormones. We often say that “the ovaries” or “the testes” secrete hormones, but it’s important to recognize that only certain cells in the gonads are responsible. The principal sex steroid in males is testosterone, which we’ve already seen is secreted by the interstitial (Leydig) cells in the testes. In females, the somatic cells of the ovarian follicles secrete feminizing steroids called estrogens. Typically, two or more specific types of estrogen molecules are secreted, but here we refer to them simply as “estrogens.” After follicles have ruptured during ovulation, the somatic cells remaining in the ovary reorganize and—in addition to estrogens—secrete another type of female sex steroid, progesterone, which is involved principally in coordinating processes associated with pregnancy. The placenta also secretes estrogens and progesterone during pregnancy.
777
LINK
For information on the roles of sex steroids during embryonic development, see Concept 35.4
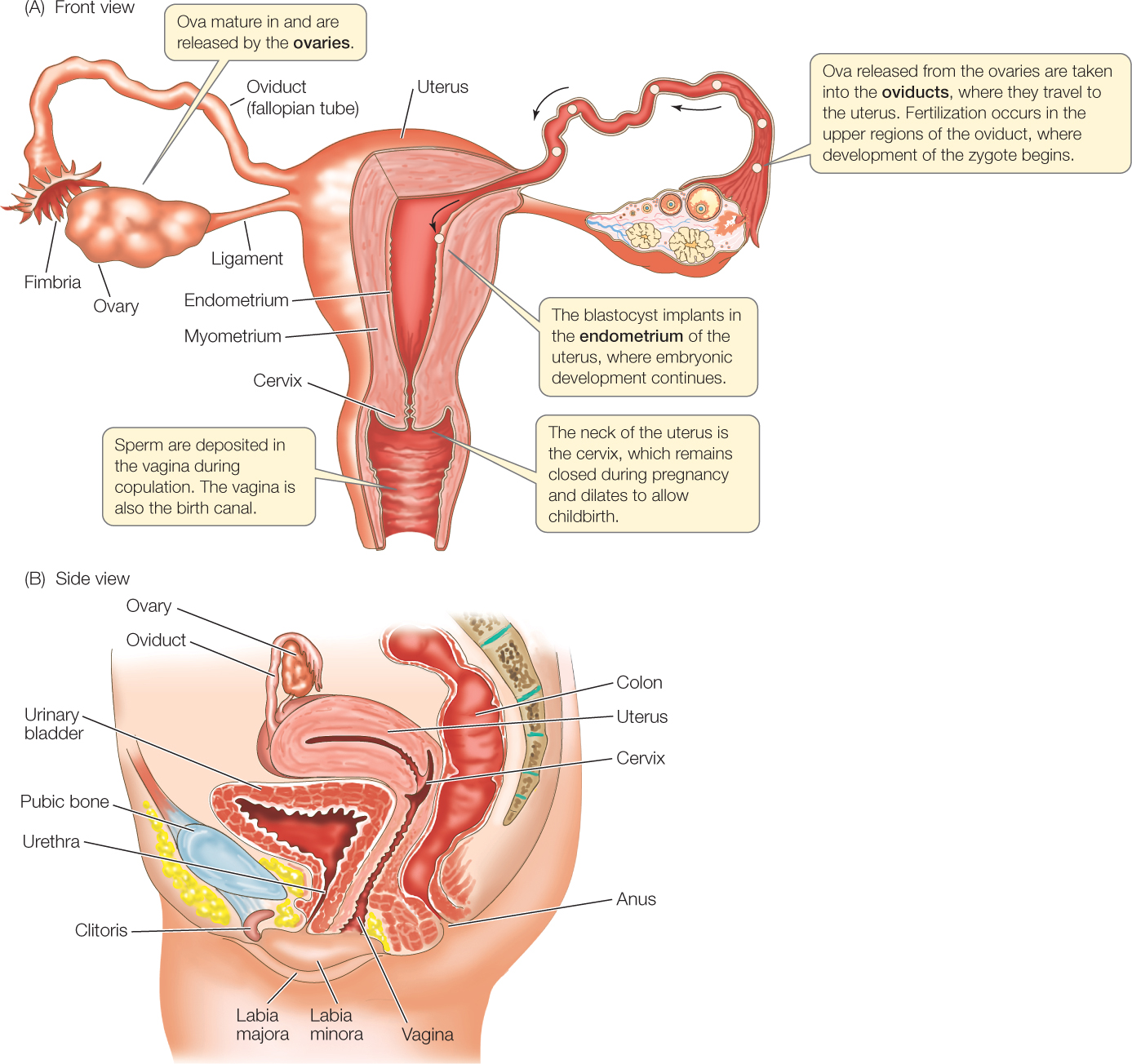
Ova mature in the ovaries and move to the uterus
The ovaries are positioned in the lower abdomen, as seen in FIGURE 37.10 (Figure 35.10 provides a larger perspective). Ova released from the ovaries make their way to the uterus by way of tubes called the oviducts or fallopian tubes. Each ovary is positioned near the funnel-shaped opening of an oviduct. When an ovum is discharged from a follicle at the surface of the ovary during ovulation, it is drawn into the oviduct, and then it is propelled toward the uterus by beating cilia and contractions of smooth muscles.
Go to ACTIVITY 37.2 The Human Female Reproductive Tract
PoL2e.com/ac37.2
Sperm are deposited in the vagina—a tubular structure that opens to the outside—during copulation (sexual intercourse). Fertilization of an ovum, if it occurs, takes place in the upper third of the oviduct. Sperm in the vagina enter the uterus at the cervix, travel through the uterus, and then travel up the oviduct to achieve fertilization. Sperm are often in the upper oviduct within 30 minutes, indicating that they are probably transported not only by their swimming powers but also by uterine contractions.
Passage of an ovum or zygote down the oviduct is relatively slow. It requires about 4 days in humans. If fertilization has occurred, the zygote undergoes its early development during this time and arrives in the uterus as an early embryo called a blastocyst (see Chapter 38). The uterus is a thick-walled organ consisting of two major parts: an outer layer of smooth muscle, the myometrium, and an inner, nurturing layer, the endometrium. The endometrium is highly vascular and composed of nutrient-providing cells, and it is where the zygote will develop further. After the blastocyst arrives in the uterus, it remains in the lumen for a few days and then buries itself in the endometrium by means of enzyme-catalyzed processes. The entry of the blastocyst into the tissue of the endometrium is termed implantation.
Ovulation is either induced or spontaneous
The immediate stimulus for ovulation is a surge in the blood concentration of LH. However, the cause of this surge differs between species that undergo induced and spontaneous ovulation.
Induced ovulation, described in our opening discussion, is ovulation triggered by copulation. Rabbits and certain cats, shrews, and camelids are induced ovulators. During copulation in an induced ovulator, the copulatory act stimulates key sensory neurons in the cervix. These neurons send action potentials to the brain, where the GnRH-secreting neurosecretory cells in the hypothalamus are stimulated. These cells release GnRH, resulting in a surge in LH secretion from the anterior pituitary into the general circulation. In rabbits that have been studied, the blood LH concentration increases in 1–2 hours to more than five times higher than usual. This LH surge causes the mature follicle to rupture.
778
APPLY THE CONCEPT: The mammalian reproductive system is hormonally controlled
In a famous and much-cited paper, M. C. Chang investigated the question: Are mammalian sperm altered by being in the female reproductive tract? He studied this question in rabbits. Because of the design of Chang’s studies, the unusual properties of rabbits we’ve mentioned, such as induced ovulation, were not of any relevance. Chang collected freshly ejaculated semen from male rabbits. He diluted the semen, and then he injected it directly into the upper third of the oviduct of fertile females at various times relative to the time that ovulated ova arrived there. The results he obtained are shown in the table.a

- What do these results indicate about the question Chang asked? Justify your answer.
- In vitro fertilization is a procedure by which ova and sperm are collected and brought together in a dish. Are Chang’s results relevant to in vitro fertilization?
aM. C. Chang. 1951. Nature 168: 697–698.
Most mammals, including humans and other primates, are spontaneous ovulators. During spontaneous ovulation, follicle rupture and release of the mature ovum are again triggered by an LH surge. However, the timing of the surge is under control of endogenous hormonal cycles in the female. “Endogenous” refers to processes that originate within an animal. The hormonal cycles in a spontaneous ovulator occur on their own rhythm, more or less independently of whether copulation takes place. We will focus on spontaneous ovulation for the rest of our discussion of females.
In most species of mammals that display spontaneous ovulation, a female ovulates in cycles, as already implied. The duration of each cycle varies among species. For instance, cycles in rats and mice are typically 4–6 days, whereas the human cycle averages 28 days. The cycles repeat, over and over, during reproductive seasons (i.e., all year in humans) unless pregnancy occurs. If pregnancy occurs, the cycles stop.
Hormones have the remarkable feature that they can affect many tissues at once, as we stressed in Chapter 35. In female reproductive cycles, hormones coordinate the ovaries, uterus, and in some cases, behavior.
Primates menstruate in each cycle that does not result in pregnancy. Menstruation is a discharge of bloody material from the uterus via the vagina. During each cycle, the endometrium undergoes preparation for pregnancy. It develops glands and abundant vascularization, and it thickens (in humans, by a factor of three to five). If pregnancy fails to occur, much of the extra growth is sloughed off, creating the bloody discharge. The cycles of the reproductive system are called menstrual cycles because of this highly visible aspect.
Mammals other than primates do not menstruate or have menstrual cycles. Often, however, they undergo dramatic changes in behavior: they cyclically enter estrus. This is the most highly visible aspect of their cycles, which accordingly are called estrous cycles. (Note the cycles are “estrous” cycles, whereas the period of sexual receptivity is “estrus.”) The endometrium cyclically prepares for pregnancy and regresses during estrous cycles, but the regression occurs by reabsorption, not sloughing off, of tissue.
Hormones secreted by the brain, anterior pituitary gland, and ovaries are important for the cycles. These organs interact and affect each other to create the cycles. If pregnancy occurs, the embryo and uterus join in.
Hormone receptors are as important as hormones. Tissue responses to hormones are highly affected by the numbers of hormone receptors present, and hormones can cause cells to modify their receptor numbers.
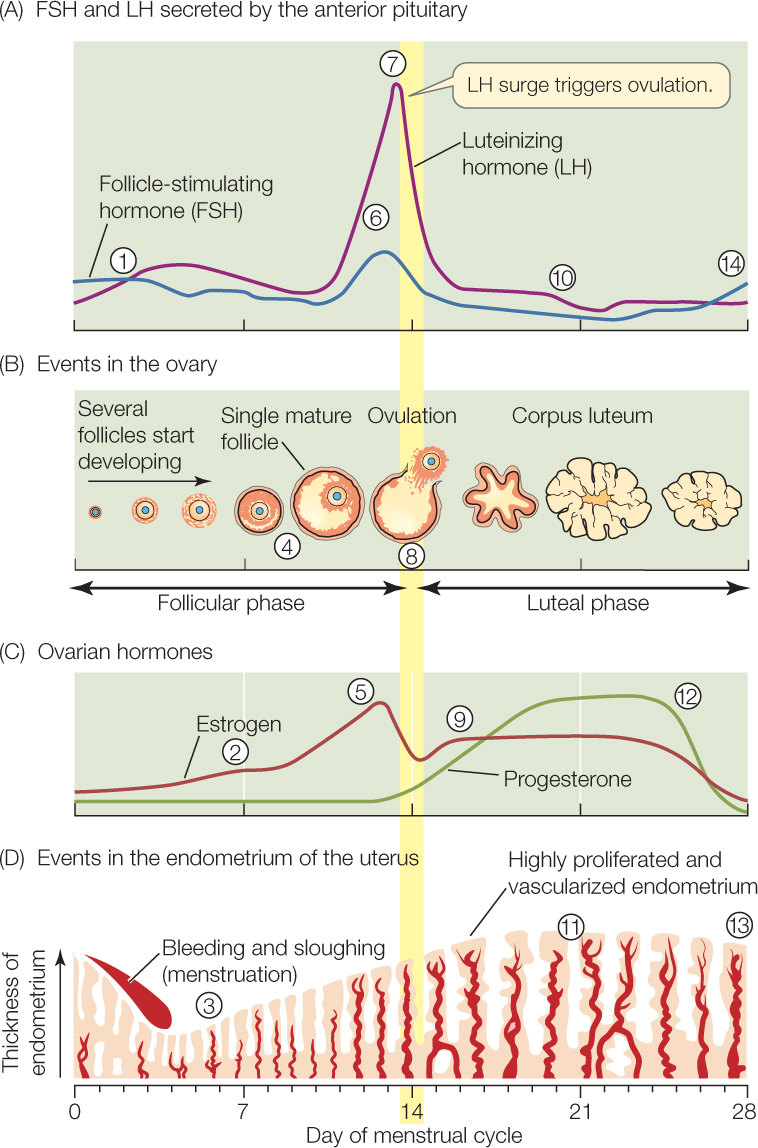
To see how cycling occurs, let’s look at the human menstrual cycle (FIGURE 37.11). By convention, each cycle is considered to start on the day menstruation begins, and that day is numbered day “0.” Follicles start their development then, and ovulation (follicular rupture) occurs on day 14. Accordingly days 0–14 are called the follicular phase of the cycle. What happens over those 14 days? The answer is complex, but by mastering it you will be rewarded with an understanding of one of the most fascinating and important processes in the human body. Follow the steps listed here by referring to the circled numbers in Figure 37.11A–D:

Go to ANIMATED TUTORIAL 37.2 The Menstrual Cycle
PoL2e.com/at37.2
 At the start, FSH and LH are secreted by the anterior pituitary at fairly high rates compared with the rates seen following ovulation during the previous cycle (compare with step 10). The FSH and LH stimulate development of follicles, including the one follicle that will mature and release its ovum. Specifically, they stimulate the somatic cells of that follicle—the granulosa and thecal cells—to secrete estrogen.
At the start, FSH and LH are secreted by the anterior pituitary at fairly high rates compared with the rates seen following ovulation during the previous cycle (compare with step 10). The FSH and LH stimulate development of follicles, including the one follicle that will mature and release its ovum. Specifically, they stimulate the somatic cells of that follicle—the granulosa and thecal cells—to secrete estrogen. Estrogen secretion gradually increases as the follicle develops.
Estrogen secretion gradually increases as the follicle develops. The increasing estrogen secretion affects the endometrium, stimulating endometrial development that prepares the uterus to accept a blastocyst if fertilization occurs. Estrogen also simulates the endometrial cells to express progesterone receptors, preparing these cells to respond to progesterone later on.
The increasing estrogen secretion affects the endometrium, stimulating endometrial development that prepares the uterus to accept a blastocyst if fertilization occurs. Estrogen also simulates the endometrial cells to express progesterone receptors, preparing these cells to respond to progesterone later on.779
 In the second half of the follicular phase, the granulosa cells of the maturing follicle increase their numbers of LH receptors.
In the second half of the follicular phase, the granulosa cells of the maturing follicle increase their numbers of LH receptors. As day 14 approaches, the blood estrogen concentration is high.
As day 14 approaches, the blood estrogen concentration is high. At this stage, estrogen stimulates the anterior pituitary and possibly the brain to bring about increasing secretion of LH and FSH.
At this stage, estrogen stimulates the anterior pituitary and possibly the brain to bring about increasing secretion of LH and FSH. Finally, estrogen stimulation of the anterior pituitary causes the surge in LH secretion.
Finally, estrogen stimulation of the anterior pituitary causes the surge in LH secretion. The LH surge has a strong effect on the follicle because of the abundance of LH receptors in the granulosa cells (recall step 4). The follicle ruptures, and ovulation occurs. In mammals that (unlike humans) undergo estrus, the peak estrogen secretion at this time stimulates the behaviors of estrus. Estrus thus coincides with ovulation.
The LH surge has a strong effect on the follicle because of the abundance of LH receptors in the granulosa cells (recall step 4). The follicle ruptures, and ovulation occurs. In mammals that (unlike humans) undergo estrus, the peak estrogen secretion at this time stimulates the behaviors of estrus. Estrus thus coincides with ovulation.
When the follicle ruptures, most of the somatic cells of the follicle are left behind in the ovary. LH (which, you’ll recall, is luteinizing hormone) stimulates those cells to reorganize to form a new hormone-producing structure called the corpus luteum (see Figure 37.11B). If pregnancy has not occurred, the corpus luteum, at this stage known as a corpus luteum of the cycle, secretes for a finite length of time: about 10 days in humans. It then starts to undergo programmed degeneration, ceasing to function entirely on day 28 of the menstrual cycle. Days 14–28 are the luteal phase of the cycle. What happens during it? Again, follow the steps by referring to the circled numbers in Figure 37.11A–D:
 The corpus luteum secretes progesterone (its principal secretion), estrogen, and a third hormone, inhibin.
The corpus luteum secretes progesterone (its principal secretion), estrogen, and a third hormone, inhibin. These hormones, acting together, suppress FSH and LH secretion by the pituitary. In primates, this suppression is so strong that it prevents new follicles from starting to develop at this stage.
These hormones, acting together, suppress FSH and LH secretion by the pituitary. In primates, this suppression is so strong that it prevents new follicles from starting to develop at this stage. Progesterone secretion from the corpus luteum has a major effect on the endometrium. Exocrine glands in the endometrium are stimulated to secrete nutrient materials, and the endometrium becomes more vascularized. These changes fully prepare the endometrium to accept a blastocyst.
Progesterone secretion from the corpus luteum has a major effect on the endometrium. Exocrine glands in the endometrium are stimulated to secrete nutrient materials, and the endometrium becomes more vascularized. These changes fully prepare the endometrium to accept a blastocyst. If fertilization has not occurred, the corpus luteum starts to undergo its programmed degeneration after about 10 days. This leads to a steep decline in progesterone and estrogen secretion.
If fertilization has not occurred, the corpus luteum starts to undergo its programmed degeneration after about 10 days. This leads to a steep decline in progesterone and estrogen secretion. Deprived of the luteal hormones (hormones secreted by the corpus luteum), the endometrium cannot maintain its highly developed state. It starts to regress, with menstruation beginning on day 0 of the next cycle.
Deprived of the luteal hormones (hormones secreted by the corpus luteum), the endometrium cannot maintain its highly developed state. It starts to regress, with menstruation beginning on day 0 of the next cycle. The disappearance of luteal hormones also sets the stage for a new cycle to begin because the luteal hormones no longer act on the pituitary or brain to suppress FSH and LH secretion. FSH and LH increase, returning the cycle to step 1, in which development of a new follicle is stimulated.
The disappearance of luteal hormones also sets the stage for a new cycle to begin because the luteal hormones no longer act on the pituitary or brain to suppress FSH and LH secretion. FSH and LH increase, returning the cycle to step 1, in which development of a new follicle is stimulated.
Pregnancy is a specialized hormonal state
If fertilization takes place, the blastocyst burrows into the tissue of the endometrium within a few days after arriving in the uterus, and there is an urgent need to maintain the endometrium in its highly developed state. In humans and many other mammals, the endometrium is prevented from deteriorating by rescue of the corpus luteum. A signal of the existence of pregnancy is sent to the corpus luteum. In response, the corpus luteum does not undergo programmed degeneration and instead becomes a corpus luteum of pregnancy, which continues to grow and secrete hormones. The signal in primates is a hormone, chorionic gonadotropin (CG), secreted by the blastocyst. In other words, the embryo emits a signal that tells the mother’s tissues to maintain the uterus in a state suitable for pregnancy. The concentration of CG rises high enough in the mother’s blood and urine to be easily detected, and many pregnancy tests work by detecting it.
780
The uterus requires progesterone to remain in a state suitable for pregnancy. In humans, after the corpus luteum is rescued, it provides adequate progesterone for 7–10 weeks. By then, the placenta—which also secretes progesterone—has become sufficiently well developed to take over.
The placenta is a structure in which maternal blood vessels and embryonic (or fetal) blood vessels are closely juxtaposed (see Figure 38.19). Thus in addition to its hormonal role, the placenta enables exchange of materials between mother and embryo. The blood of the mother and that of the embryo do not mix. However, nutrients and O2 move across vessel walls from the maternal blood into the blood of the embryo, and CO2 and other wastes move from the embryo into the maternal blood.
At the end of pregnancy, still another hormone, oxytocin (secreted by the hypothalamus and posterior pituitary gland; see Concept 35.3 and Figure 35.6) plays an essential role. As we have already discussed (see Figure 29.17), oxytocin stimulates the uterine muscle layer (the myometrium) to contract with ever-increasing force during the birth process so that the baby is expelled from the uterus. Following birth, oxytocin and the anterior pituitary hormone prolactin stimulate the mammary glands (breasts) to produce and eject milk.
Male sex organs produce and deliver semen
The testes of most mammals are located outside the body cavity in a sac of skin called the scrotum (see Figure 37.3). During early development they are inside the abdomen in the same positions as the ovaries, but they migrate into the scrotum as development proceeds (see Concept 35.4). In human males, the testes are normally in the scrotum all the time. In many other species, such as certain mice, the testes are in the scrotum only during the reproductive season, being withdrawn into the abdomen at other times. The testes of most mammals need to be about 2°C cooler than ordinary abdominal temperature (37°C) for normal production of viable sperm, and being in the scrotum permits this.
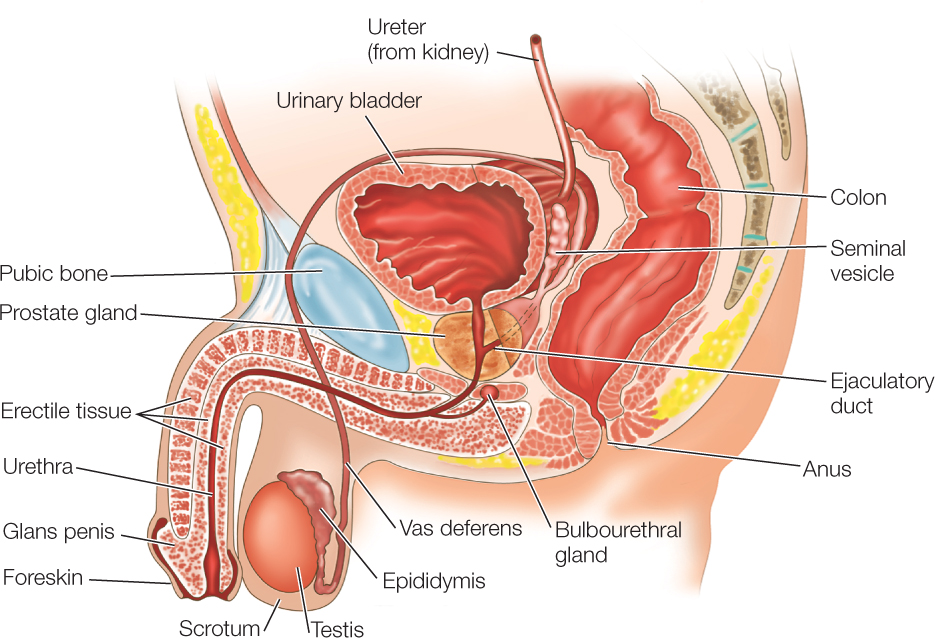
After sperm are produced in a testis, they are stored in the epididymis and vas deferens. The epididymis is a convoluted tubular structure located next to the testis in the scrotum (see Figure 37.5). The vas deferens is the tube that leads from the epididymis to the urethra, the passage in the penis through which urine and semen flow to the outside (see Figure 37.3). The vas deferens and urethra are endowed with smooth muscle, which contracts to propel semen during ejaculation.
Semen, the fluid expelled from the penis during ejaculation, consists of a mix of sperm and fluids secreted by male accessory reproductive glands. In humans, the glands that provide most of the fluids in semen are the paired seminal vesicles and the single prostate gland (FIGURE 37.12). Sperm are mixed with these fluids as ejaculation occurs. The fluids provide a supportive medium for the sperm, including fructose and other energy sources. The paired bulbourethral glands secrete a clear mucus prior to ejaculation. This fluid often emerges from the penis and may contain sperm even though it is not semen.
Go to ACTIVITY 37.3 The Human Male Reproductive Tract
PoL2e.com/ac37.3
The shaft of the human penis is filled almost entirely by three compartments of spongy tissue that can be expanded with blood (see Figure 37.12). Erection occurs when these compartments are inflated with blood at high pressure until the penis is stiff. Nitric oxide (NO) is the agent that immediately controls erection. When NO is released from parasympathetic nerve endings in the penis, it leads to dilation of blood vessels that permit blood to fill the spongy tissues. NO is short-lived. It acts by increasing synthesis of cyclic GMP (cyclic guanosine monophosphate), which in turn acts as a second messenger (see Concept 5.6). Medications for erectile dysfunction (impotence) inhibit an enzyme that breaks down cyclic GMP. This permits the second messenger to act longer.
Many mammals have a bone in the penis that helps stiffen it for copulation. Included are dogs, seals, rodents, and some primates (but not humans).
Hormones play as crucial a role in male reproductive function as in female. Reproductive processes occur in far more steady ways in males than females, however, and the controlling hormones do not cycle in the dramatic ways seen in females.
781
Testosterone is required for spermatogenesis. In human males, the interstitial cells secrete an abundance of testosterone during a period of fetal life and again during the first year after birth. After that, testosterone levels remain low until puberty begins. At puberty, the blood testosterone level increases dramatically, and thereafter it stays high for the rest of life, although it declines gradually after middle age. Spermatogenesis begins at puberty when the testes mature and thereafter continues without interruption throughout life. Many nonhuman mammals exhibit strongly seasonal reproduction. In these animals, testosterone secretion and spermatogenesis often occur only in reproductive seasons. In those seasons, however, the animals exhibit the same steadiness as seen in humans in testosterone secretion and spermatogenesis.
The pituitary hormones FSH and LH play central roles in males. LH stimulates testosterone secretion by the interstitial cells. In this way it plays a mandatory role in spermatogenesis. FSH and testosterone, acting together, stimulate the Sertoli cells to support spermatogenesis.
Many contraceptive methods are available
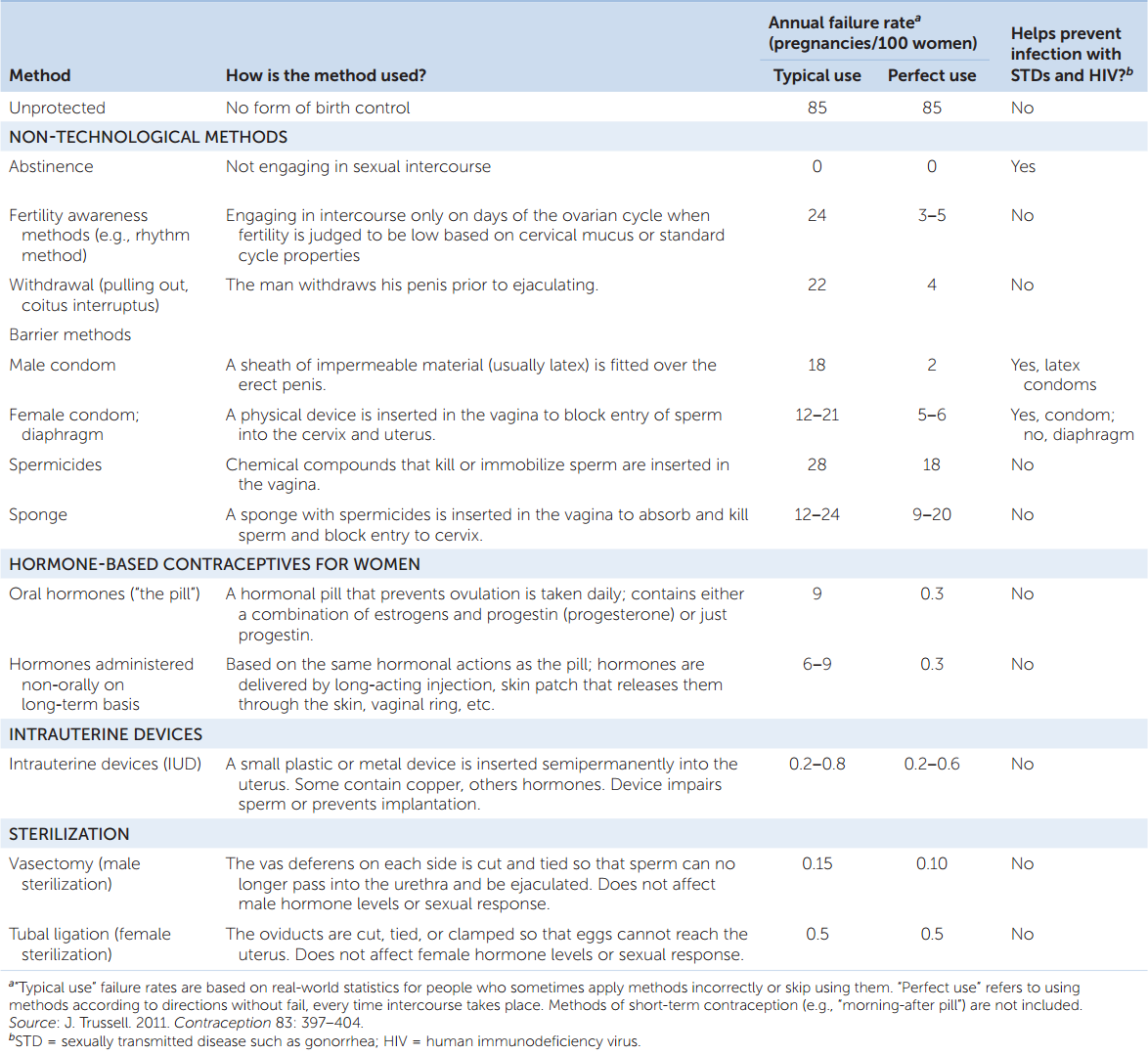
Useful contraceptive methods are based on an accurate understanding of the biology of reproduction. They fall into a several categories (TABLE 37.1). The simplest to understand is abstinence. Without sperm in the female reproductive tract, pregnancy cannot occur. Several methods are based on blocking sperm from entering the uterus during and after sexual intercourse. This is the rationale for condoms (male and female) and diaphragms. Some methods, such as birth control pills, manipulate a woman’s hormone cycle so that ovulation does not occur.
782
Table 37.1 presents contraceptive failure rates in the United States according to a recent assessment. In many ways, the most informative aspect of the data is the comparison of failure rates for “typical” and “perfect” applications of the methods. Notice the large differences for some methods. For example, latex male condoms are estimated to have an annual failure rate of 2 pregnancies per 100 women (whose partners use condoms) when the condoms are used exactly according to directions. However, among people in the general population who say they use male condoms as their method of contraception, the failure rate is almost 10 times greater. This difference reflects deficiencies in how and when condoms are put on and removed, and it also reflects instances when avowed condom users skip using them. The message is clear: many methods are not very effective unless the users follow the directions in detail and without fail.
CHECKpoint CONCEPT 37.2
- What is the role of chorionic gonadotropin (CG) in human reproduction, and why is it a target for pregnancy tests?
- Where do the fluids come from that are expelled from a man’s penis before and during ejaculation?
- Taking specific account of FSH, explain the relationship between the cycling of estrogen secretion and the cycling of follicle development during the human menstrual cycle.
In countless ways, reproduction must be integrated with the rest of an animal’s life cycle. If reproduction requires nutrients, for example, it must be integrated with processes that provide nutrients. Thus a final aspect of reproduction is how it relates to other aspects of the life cycles of animals.