CONCEPT38.3 Gastrulation Produces a Second, then a Third Germ Layer
Gastrulation involves major movements of cells from the external surface to the interior of the embryo. These cell movements create a three-layered body plan and set the stage for development of the first organs. Triploblastic animals (see Concept 23.1) create all of their organs and tissues from three basic germ layers—ectoderm, endoderm, and mesoderm. These layers are formed by the process of gastrulation. The resulting embryo is known as a gastrula.
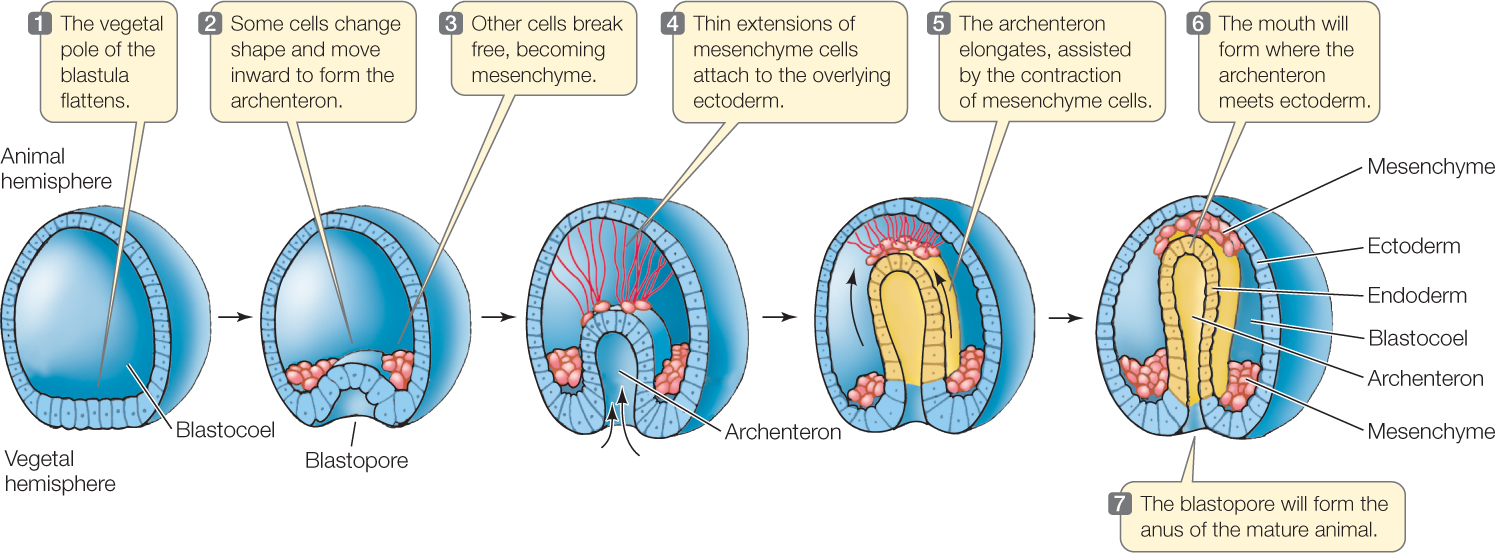
Just as the amount of yolk affects cleavage, it also affects how gastrulation occurs. The process is easiest to understand in radially cleaving eggs with little yolk, such as the much-studied eggs of sea urchins (FIGURE 38.9). One wall of the blastula (the vegetal pole) bulges inward, or invaginates, as if someone were poking a finger into a hollow ball. The new space that forms is the archenteron, or “first gut.” The opening into the archenteron is the blastopore. Recall that in deuterostomes, the blastopore becomes the anus and the mouth (the second opening) forms opposite it. The outer layer of cells is now ectoderm, and the wall of the archenteron is endoderm and future mesoderm. The archenteron elongates, assisted by contractions of wandering cells called mesenchyme cells. The archenteron will eventually squeeze the blastocoel out of existence.
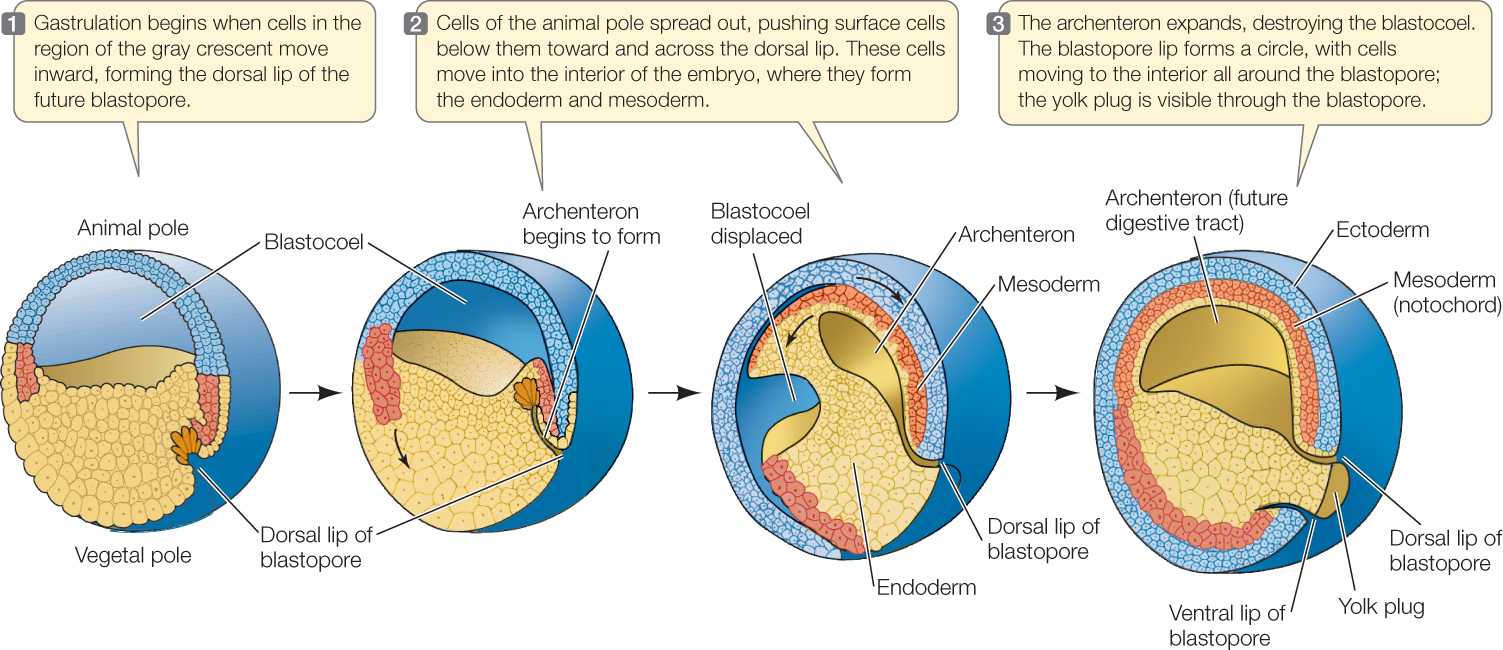
In eggs such as those of amphibians, which contain a moderate amount of yolk, cleavage continues to produce many ever-smaller cells in the animal hemisphere and larger cells in the vegetal hemisphere. To initiate gastrulation, some cells in the gray crescent invaginate to form a slitlike blastopore (FIGURE 38.10). Small blastomeres of the animal pole then begin to roll like a sheet over the dorsal lip of the blastopore and push into the blastocoel. At the same time, cell divisions continue to increase the number of cells. Because the small cells of the animal hemisphere continue to divide more rapidly than the larger cells of the vegetal hemisphere, these smaller cells grow over and surround the larger cells of the vegetal hemisphere. As these smaller cells reach the blastopore, they too move inward so that the inward migration occurs all around the blastopore. These cells will form much of the endoderm and mesoderm of the developing embryo. The blastopore becomes the anus.
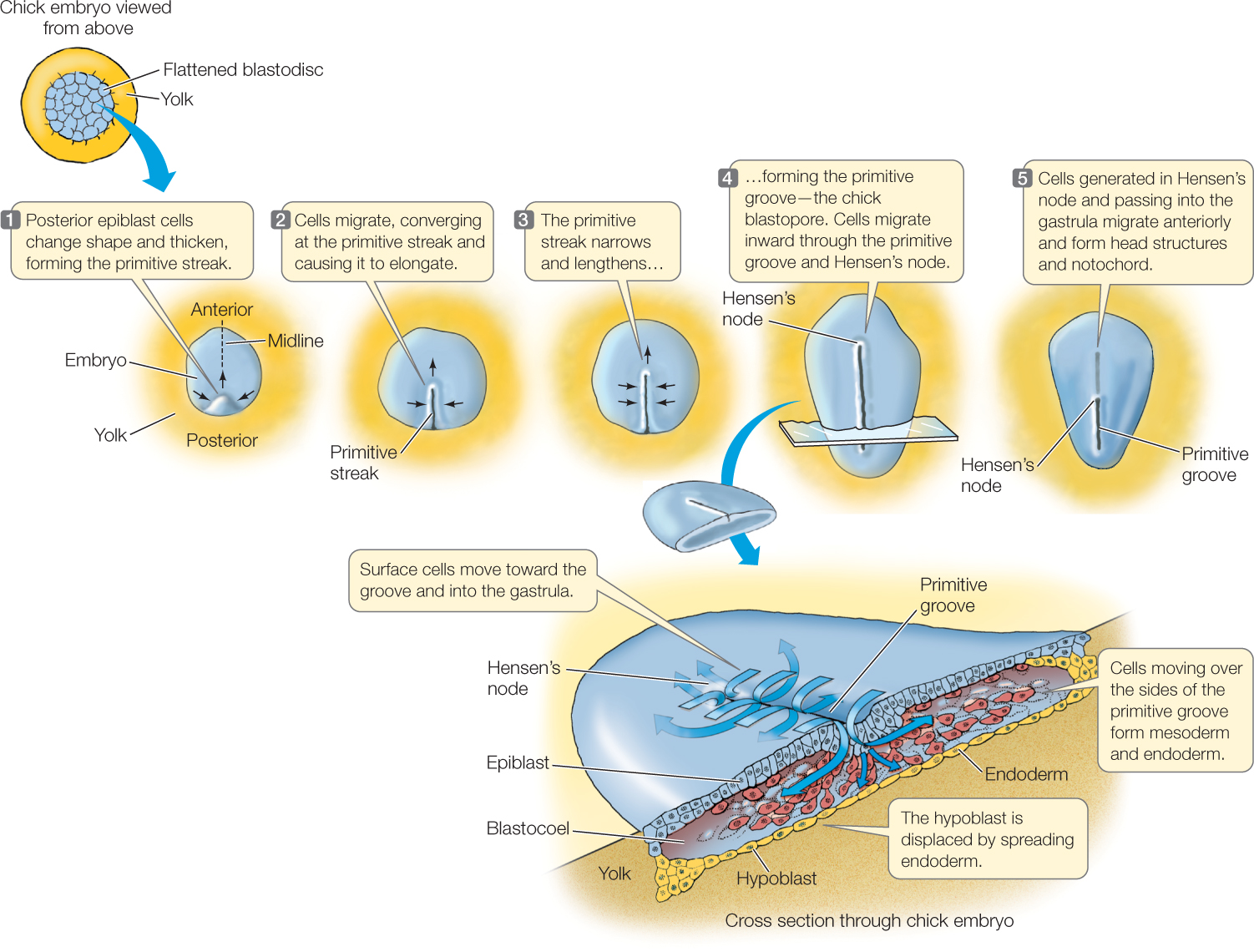
In very yolky eggs like those of birds, in which the blastodisc forms on top of the yolk, the blastopore forms as a groove in the primitive streak, a collection of cells marked at the anterior end by Hensen’s node (FIGURE 38.11). Cells that will become mesoderm and endoderm migrate inward through Hensen’s node and all along the groove of the primitive streak.
Placental mammals retain the avian/reptilian gastrulation pattern, even though they lack yolk. Mammals and reptiles, including birds, are all amniotes (i.e., they produce eggs that develop extraembryonic membranes), so it is not surprising that they share certain patterns of early development. In Concept 38.2 we discussed the development of the mammalian inner cell mass and the outer trophoblast. As in avian development, the inner cell mass of placental mammals splits into an upper layer called the epiblast and a lower layer called the hypoblast (see Figure 38.8). The embryo forms from the epiblast, while the hypoblast contributes to the extraembryonic membranes that will encase the developing embryo and help form the placenta, the temporary organ that nourishes most mammalian embryos. Gastrulation occurs in the mammalian epiblast as it does in the avian epiblast. A primitive groove forms, and epiblast cells migrate through the groove to become layers of endoderm and mesoderm.
All three types of gastrulation described above can be found among the protostomes, but recall that there is a major difference in the fate of the blastopore. In protostomes, the blastopore (the first opening) becomes the mouth and the anus forms at some distance away; in deuterostomes, the blastopore becomes the anus, and the mouth breaks through secondarily. This is why animals are called protostomes (“mouth first”) or deuterostomes (“mouth second”).
LINK
For more on the differences between protostomes and deuterostomes, see Concept 23.1
Gastrulation results in the production of the three germ layers and sets the stage for the formation of the coelom, a space completely enclosed in mesoderm. Your body cavity—the space between the gut and body wall—is a coelom (see Concept 23.1). Because of the coelom, gut contractions and heartbeats are independent of an animal’s outer body wall. To understand the formation of the coelom, it is best to consider the mesoderm and coelom together.
In deuterostomes, the mesoderm often forms from the archenteron wall, and the coelom develops within in it, sometimes as a direct out-pouching of the archenteron (FIGURE 38.12A). This type of mesoderm and coelom formation is believed to be characteristic of deuterostomes, although it may be highly modified by yolk and other factors.
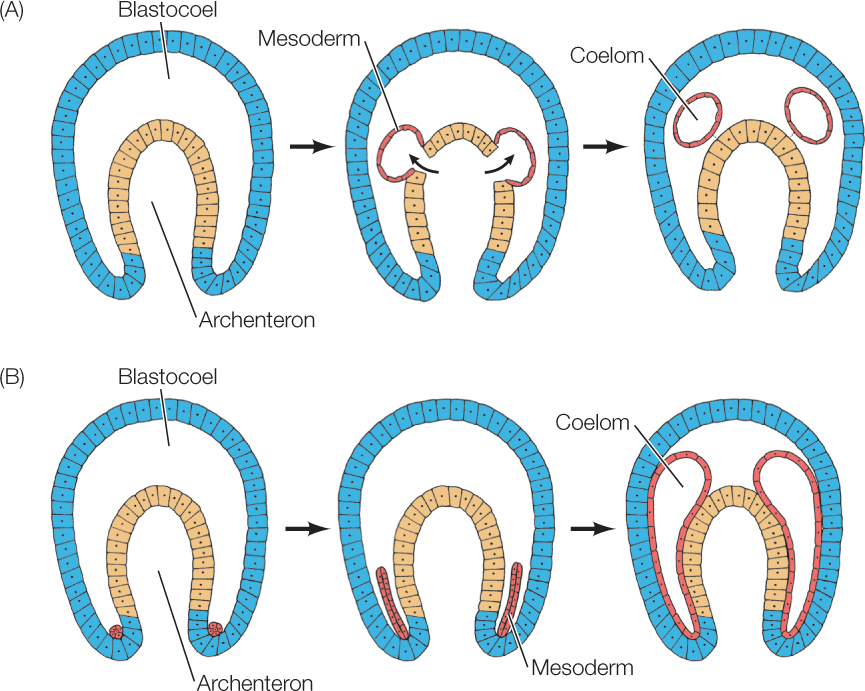
Protostomes show a different pattern that fits with the fact that specific blastomeres usually give rise to each tissue and organ (mosaic development). For example, in the snail Ilyanassa, one recognizable small blastomere, 4d, forms the bulk of the mesoderm. This single cell (4d) received information directing production of mesoderm from the polar lobe (see Figure 38.4), undergoes divisions and forms most of the mesoderm. The coelom forms as a split in the prospective mesoderm cells (FIGURE 38.12B).
Whatever the mechanism of gastrulation, the process produces an embryo with three germ layers—ectoderm, endoderm and mesoderm—that are positioned to influence one another through tissue interactions.
CHECKpointCONCEPT38.3
- What does the blastopore become in protostomes? What does it become in deuterostomes?
- What are the major similarities and differences between sea urchin gastrulation and frog gastrulation?
- Why is the primitive streak in the chick embryo the equivalent of the blastopore?
- Contrast mesoderm formation in the non-yolky eggs of a deuterostome such as a starfish with those of a protostome such as a snail. Compare coelom formation in these two groups.
Following gastrulation and the subsequent formation of three germ layers, an embryo is ready to start forming the organs of the new individual. Let’s now look at that key process.