Concept 40.6: Behavior Helps Structure Ecological Communities and Processes
In ecological communities, the behaviors of animals often give critically important structure to the use of time and space and to interrelationships among species. One way that behavior has this effect is the structuring of the 24-hour day into activity periods. Some animals are active only during daylight hours. Others are active only at night. These behavioral differences can determine whether two species encounter each other. For example, because insect-eating bats are typically nocturnal, spending their days in secluded places such as caves and attics, diurnal insects are not threatened by the bats even though the two live in the same ecological community.
Behavior helps maintain species
In much of North America, the principal native mice are members of the genus Peromyscus, and it is common for two species in this genus to be found living in one general region. White-footed mice (P. leucopus) and cotton mice (P. gossypinus) are one example. When males and females of these two species are housed together in cages, they breed readily and produce fertile offspring. In nature, however, hybrids are rarely found. To a large degree, behavior explains this difference. In nature the two species prefer different types of woodlands. They thus live in different places and probably rarely meet. We can also hypothesize that if they were to meet, they would detect oddities in each other’s interactive behavior and separate before mating. Many similar cases are known of species that can be demonstrated to be interfertile yet remain distinct in undisturbed natural habitats. Their behaviors keep them apart in such habitats. Behavior is thus a key factor in allowing species to maintain their species distinctions.
The species of Darwin’s finches in the Galápagos Islands are of particular interest because intense study of the finches has provided unusually detailed insight into hybridization in the wild. Peter and Rosemary Grant have established by analyses of DNA that, among the several species of ground-living Darwin’s finches, about 1 percent of birds that grow to adulthood in the wild are fertile hybrids of two distinct species. The hybridization that is observed demonstrates that the species of Darwin’s finches are not genetically walled off from each other. Instead, behavior ordinarily keeps the species distinct. Males ordinarily learn their songs from their fathers (see Concept 40.2), and a female ordinarily will not mate with a male singing a song other than the correct one for her species. These behaviors prevent most cross-breeding.
LINK
Behaviors that prevent cross-breeding are an important mechanism of reproductive isolation and speciation; see Concept 17.4
Animals often behaviorally partition space into territories or home ranges
In many types of animals—including ones as diverse as fish, dragonflies, and birds—individuals within a single population restrict their movements to limited areas. For example, FIGURE 40.16 shows use of space by six adult male white-footed mice within a 4-hectare forest. Each mouse roamed within a defined area on the forest floor, with almost no overlap among the areas occupied by the different mice.
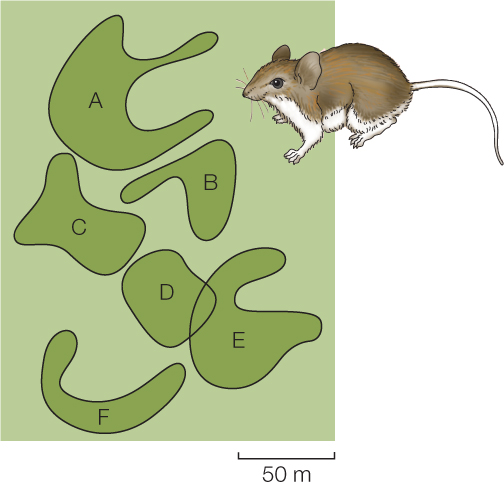
In cases like this, behavior acts to structure the space available. Rather than interacting ecologically with the entire forest, each individual interacts with the ecological community in just a small part, which is more or less distinct from the parts with which other, neighboring individuals are engaged.
842
The region occupied by an individual is known as its territory if the individual actively keeps out other individuals of the same species. A region is known as an individual’s home range if other individuals are not excluded. Home ranges often overlap considerably. Both territories and home ranges provide familiarity. An individual occupying a territory or a home range learns all the locations within its living area where it can escape attacks or find resources or shelter. When an individual defends a territory, it enjoys the additional benefit of having sole use of any resources present.

Go to ANIMATED TUTORIAL 40.3 The Costs of Defending a Territory
PoL2e.com/at40.3
Behavior helps structure relationships among species
For understanding behavioral relationships among species in ecosystems, ecologists often use a cost–benefit approach. Such an approach assumes that an animal has only a limited amount of time and energy, and therefore—in its relationships with other species—cannot afford to engage in behaviors that cost more to perform than they bring in benefits. In cost–benefit analysis, ecologists quantify both costs and benefits. Then, by calculating relative costs and benefits, they make—and test—predictions about which behaviors to expect.
A specific case studied in detail by Bernd Heinrich is the choice of flowers in the foraging ecology of bumblebees (Bombus). As a bumblebee forages in a flower patch, it flies from flower to flower, landing on flowers to collect nectar (FIGURE 40.17). During this process, the bee must keep its flight muscles warm so it will be able to fly. Keeping them warm during flight is no problem because the working muscles produce heat at a high rate. Keeping them warm when the bee has landed on a flower requires extra heat production when the air is cold for the same reason a house furnace needs to make heat at an increased rate in cold weather. Adding all these considerations together, Heinrich calculated the energy cost per minute of foraging and determined that the cost is considerably greater in cold air than in warm air.

Heinrich also calculated the benefit of foraging. To do this, he measured how much nectar could be obtained from each flower and the sugar concentration of the nectar. Then he calculated the energy a bee could obtain from each flower by metabolically oxidizing the sugar in the flower’s nectar.
One common flowering plant in Heinrich’s study area was a rhododendron with large flowers. A bumblebee got 1.7 joules (J) of energy by taking the nectar from each rhododendron flower. In cold air at 0°C, a bee’s cost of foraging was about 12.5 J per minute. Thus a bumblebee needed to harvest the nectar from only about 7 flowers each minute for its foraging benefit to equal its foraging cost. Actually, a bee could harvest 20 flowers per minute, receiving a benefit that far exceeded its cost—and enabling the bee to take energy back to its hive. Heinrich predicted that bumblebees would forage on rhododendron flowers in both cold and warm weather, and this is what the bees do.
Another common plant was wild cherry. Heinrich noticed that bumblebees foraged on the cherry flowers only in warm weather. By applying his cost–benefit analysis, he discovered why. Cherry flowers are smaller than rhododendron flowers and provide only 0.2 J of energy each. Accordingly, at 0°C a bee would have to harvest more than 60 cherry flowers per minute just to get enough nectar to equal its cost of foraging. Bumblebees cannot harvest that many flowers per minute. Thus they would have to go into an energy deficit—spending more energy than they received—to forage on cherry flowers in 0°C air.
With this study, Heinrich showed that bumblebee foraging behavior is structured in ways that keep a bee in energy balance. This fact, in turn, structures the relationship between bumblebees and the plants that flower in their ecosystems.

Go to ANIMATED TUTORIAL 40.4 Foraging Behavior
PoL2e.com/at40.4
LINK
A cost–benefit approach can also be used to analyze life history trade-offs during evolution; see Concept 42.3
CHECKpoint CONCEPT 40.6
- How might Lee Metzgar’s study (see Concept 40.2, Figure 40.4) on the survival of certain white-footed mice in the presence of screech owls help explain the existence of home range behavior?
- Imagine that on a cold day a bee requires 15 joules of energy per minute to forage continuously. If the bee can harvest 20 flowers of a certain species every minute, and gets 0.8 J of energy for each flower it visits, would it be to the bee’s energetic benefit to forage on those flowers that day?
- Animals that actively defend a territory gain sole access to all the energy sources available in that territory, a major energetic benefit. What might some of the energetic costs of territory maintenance be?
843
Question 40.2
In what ways might schooling behavior and pairing behavior be advantageous for the individuals involved?
ANSWER Schooling behavior can be advantageous in several ways. For example, a school has more sense organs than a single individual. For this reason, a school may detect predators sooner than an individual so that evasive action can be taken. Similarly, a school may be able to find valuable resources in the environment more rapidly or efficiently. Pairing behavior could have several advantages—one being that in male–female pairs, each individual is assured of a mate.