Concept 44.4: Diversity Patterns Provide Clues to What Determines Diversity
Species diversity is not uniform around the world. Geographic patterns in diversity shed light on factors that affect diversity by modifying the balance between processes that add species to and remove species from communities.
Species richness varies with latitude
When the German naturalist Alexander von Humboldt traveled around Central and South America, Europe, and Asia two centuries ago, he noted that “the nearer we approach the tropics, the greater the…variety of structure…of organic life.” This recognition that species richness increases toward the equator has been amply confirmed since then. A 2-hectare plot of tropical forest in Malaysia, for example, can contain 227 tree species, whereas an equivalent plot of temperate forest in Michigan contains only 10 to 15. There are 56 bird species that breed in Greenland, 105 in New York State, 469 in Guatemala, and 1,395 in Colombia. And although some taxa appear not to follow the pattern—of some 20,000 known bee species on Earth, for instance, about half are found in temperate regions and half in the tropics—many other taxa do (FIGURE 44.9).
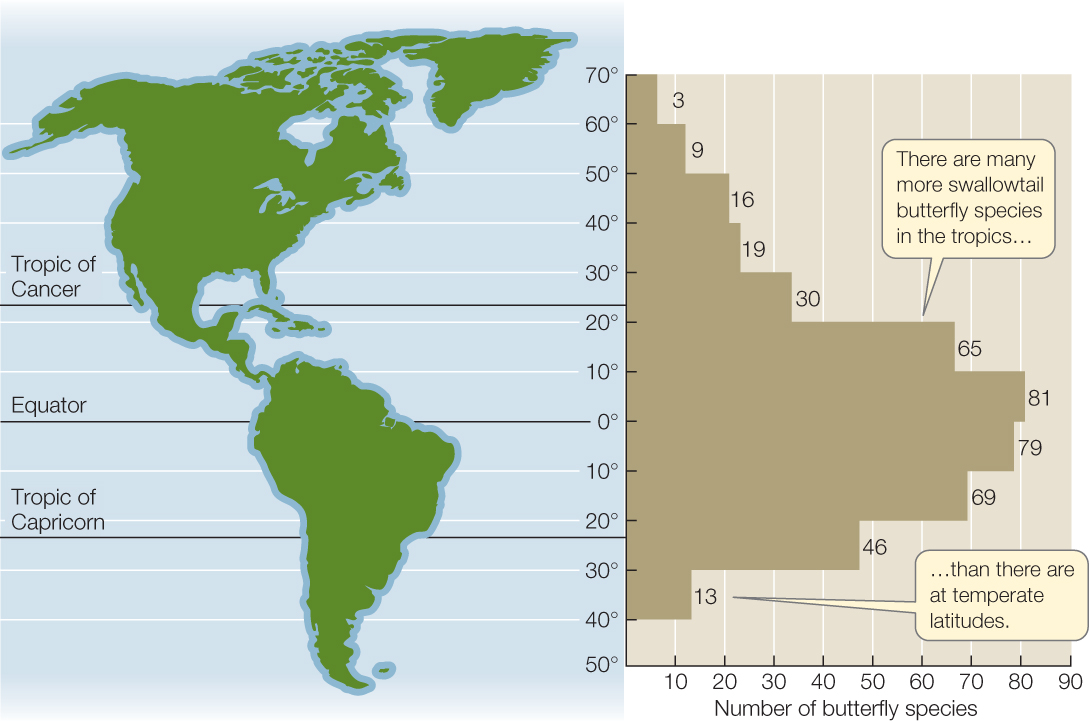
What is it about the tropics that supports such diversity? Over long time spans, climate conditions have been more stable in the tropics than in temperate regions. The tropics were never subjected to the dramatic glacial advances that caused massive shifts in the geographic ranges of species at higher latitudes and led to many extinctions. Tropical communities are certainly dynamic, and they are strongly influenced by disturbances (such as the death and fall of individual trees in rainforests), but the absence of disturbance at large spatial scales may have allowed these communities to retain more of their species than temperate communities, leaving them with higher species richness today.
We also know that climate changes with latitude. The tropics receive abundant solar energy input, which makes them warm and wet (see Concept 41.2). These conditions promote rapid growth of primary producers, so NPP, like diversity, increases toward the equator. High NPP can enhance diversity (as well as be enhanced by diversity; see Concept 44.3) if greater energy input into food webs allows species to maintain large population sizes, even if they are more specialized in their use of resources or habitats. In this way, greater energy flow through tropical communities could facilitate the coexistence of a greater number of species with narrow, specialized niches.
Variation in habitat structure could interact with greater niche specialization to amplify tropical diversity. In general, diversity is higher in more structurally complex habitats. Bird species diversity, for example, increases with the diversity of foliage height in deciduous forests. If tropical species have more specialized habitat requirements, and if tropical vegetation is more structurally complex by virtue of greater plant species richness, then complex habitat structure and productivity may interact to increase tropical diversity.
907
Ecologists still have much to learn about the relative contributions of these various factors to the puzzle of tropical diversity, but the research stimulated by that puzzle has demonstrated that disturbance patterns, productivity, and habitat structure all affect species diversity.
Species richness varies with the size and isolation of oceanic islands
The numbers of species found on oceanic islands exhibit regular patterns that have long fascinated ecologists (FIGURE 44.10). First of all, species richness is greater on large than on small islands. And second, species richness is greater on islands near a mainland than on more distant islands. What can explain these differences?
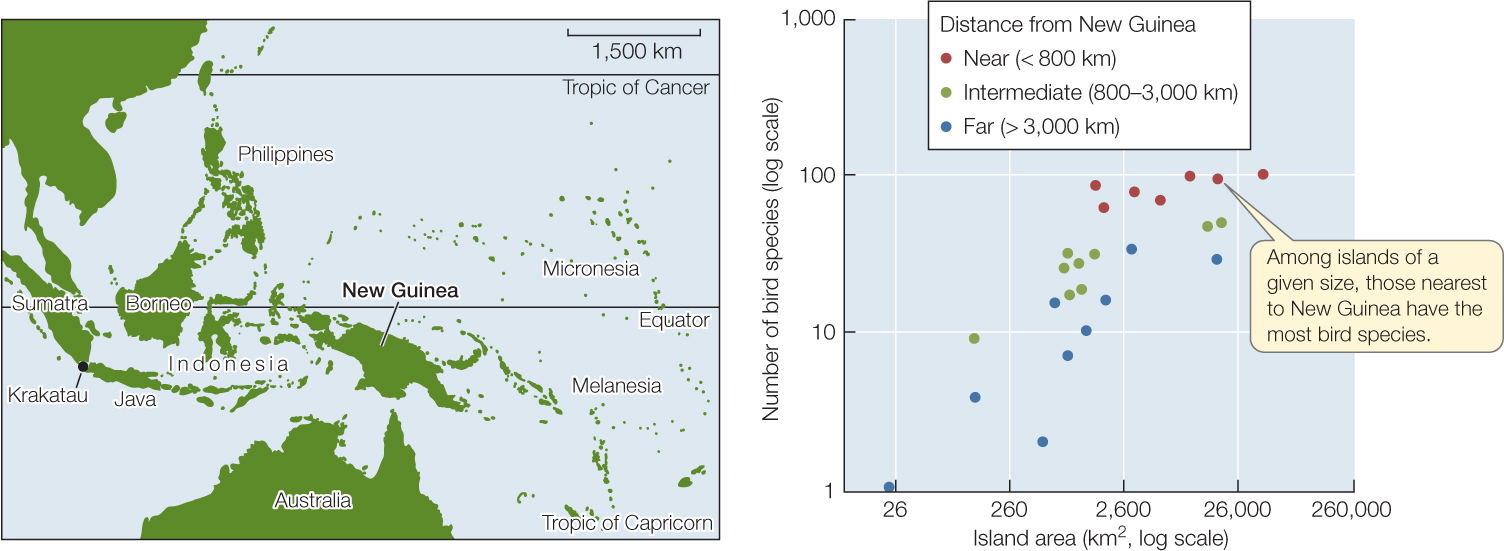
As we saw in the case of Krakatau, communities are assembled by colonization, along with occasional extinctions. Just as a population reaches an equilibrium size when additions (births) and losses (deaths) are equal, it seems plausible that species richness on an island might ultimately represent an equilibrium between the rate at which new species colonize and the rate at which resident species go extinct. This idea forms the basis for the theory of island biogeography, proposed in 1963 by Robert H. MacArthur and Edward O. Wilson. The theory can be summarized as follows:
908
- An oceanic island gains species only if they colonize from elsewhere—usually from nearby islands or continents, whose communities make up the species pool of potential colonists.
- The rate at which new species arrive on an island declines as the island fills with species. At first, every immigrant represents a species new to the island, but as the island fills up, more and more of the arriving individuals represent species already resident there. (If all species in the mainland species pool were present on the island, the colonization rate would be zero.)
- The overall rate at which species are lost from the island—the extinction rate—increases as the island fills with species. In any time period, each species has some chance of going extinct from various causes, and as the number of species increases, so does the overall number of species that go extinct per unit of time.
- The number of species on an island—its species richness—stops changing if the colonization rate equals the extinction rate. When we plot colonization and extinction rates on a graph, this equilibrium species richness is represented by the point at which the two lines cross, Ŝ (FIGURE 44.11A).
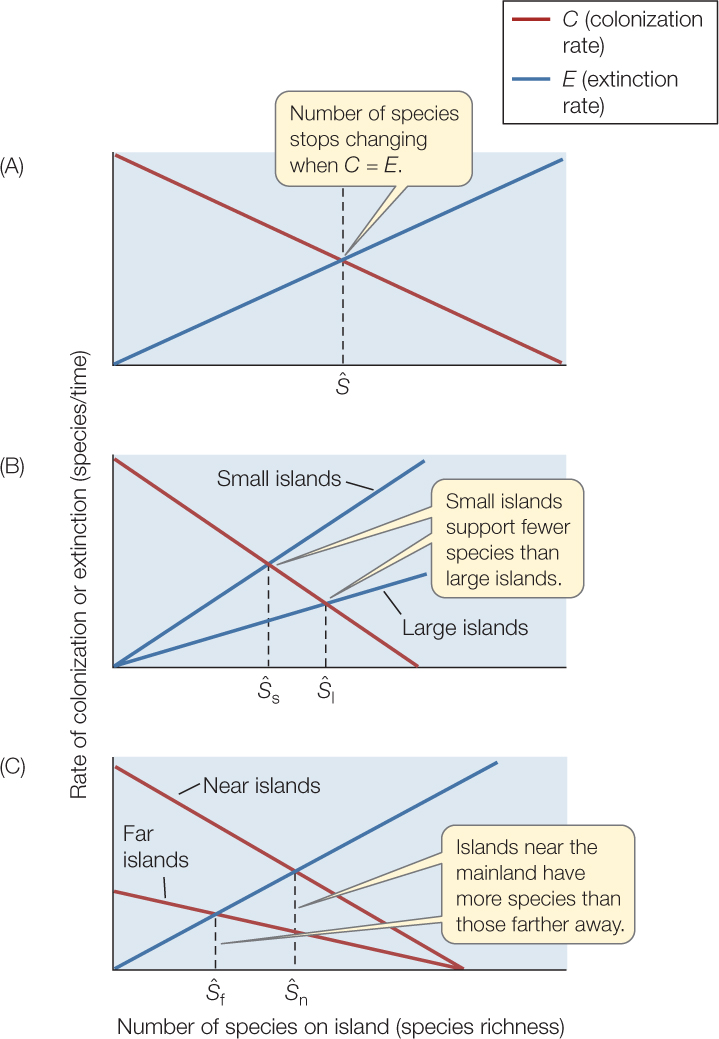 Figure 44.11: MacArthur and Wilson’s Theory of Island Biogeography (A) Species richness reaches a stable equilibrium (Ŝ) when the colonization rate equals the extinction rate. (B) Smaller islands have larger per-species extinction rates, and hence lower equilibrium species richness. (C) Islands more distant from the mainland have lower colonization rates, and hence lower equilibrium species richness.
Figure 44.11: MacArthur and Wilson’s Theory of Island Biogeography (A) Species richness reaches a stable equilibrium (Ŝ) when the colonization rate equals the extinction rate. (B) Smaller islands have larger per-species extinction rates, and hence lower equilibrium species richness. (C) Islands more distant from the mainland have lower colonization rates, and hence lower equilibrium species richness. - Population sizes will decline as island size decreases, and since small populations are more at risk of extinction, the extinction rate for small islands should rise more steeply than that of large islands as a function of total species on the islands. Thus the equilibrium species richness should be greater on large than on small islands (FIGURE 44.11B).
- Similarly, fewer wind- and water-borne seeds, and fewer dispersing animals, will encounter a distant island than one closer to the mainland species pool. As a result, the colonization rate, and thus the equilibrium species richness, will be higher on close islands than on distant ones (FIGURE 44.11C).
The theory of island biogeography successfully explains the patterns of species richness found not only on oceanic islands, but also on “islands” of one habitat type surrounded by a “sea” of a different habitat type. It has proved to be one of the most successful explanatory theories in ecology (FIGURE 44.12).
Investigation
HYPOTHESIS
The number of species on an island before it is experimentally defaunated will decrease with distance from a mainland source of colonists, and this number will be reestablished after the island has been recolonized.
METHOD
- Take a census of the terrestrial arthropod species on four small mangrove islands of equal size (11–12 m diameter) but at different distances from a mainland source of colonists.
- Erect scaffolding and tent the islands. Fumigate with methyl bromide (a chemical that kills arthropods but does not harm plants).

- Remove tenting. Monitor recolonization for the following 2 years, periodically taking a census of arthropod species.
RESULTS
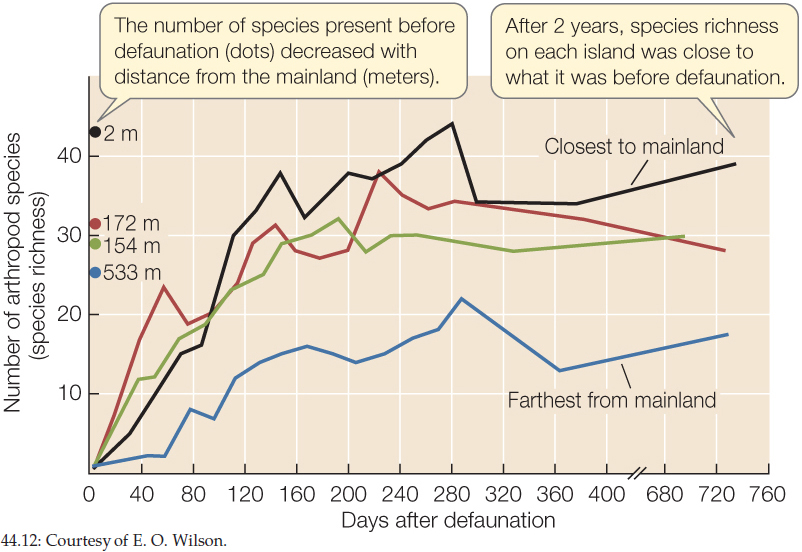
The number of species present before defaunation decreased with distance from the mainland. Two years after defaunation, each island had about the same number of species it had before the experiment.
CONCLUSION
The results support predictions of island biogeography theory.
Go to LaunchPad for discussion and relevant links for all INVESTIGATION figures.
aD. S. Simberloff and E. O. Wilson. 1969. Ecology 51: 934–937.

Go to ANIMATED TUTORIAL 44.2 Island Biogeography Simulation
PoL2e.com/at44.2
CHECKpoint CONCEPT 44.4
- Describe three factors that may contribute to the high species richness of tropical rainforests.
- Tropical coral reef communities are among the most species-rich of any on Earth. Speculate on factors that might explain their diversity.
- What do Figures 44.11B and C tell you about the predicted rate of change in species composition (species turnover rate) at equilibrium on small versus large islands and on islands distant from versus close to a mainland? (Hint: think of turnover rate as the number of new species that replace previous species in a period of time.)
Quantitative studies of species richness and the theory of island biogeography have contributed greatly to our knowledge of the structure and function of ecological communities. We will now turn to some ways in which this knowledge can be applied to conserving and restoring these communities.
909