10.4 CELL ADHESION
The cell, as we know, is the fundamental unit of living organisms (Chapter 1). Some conservative estimates place the number of cells in an adult human being at between 50 and 75 trillion, whereas others place the number at well above 100 trillion. Whichever estimate is more accurate, humans, as well as all complex multicellular organisms, are made up of a lot of cells! What then keeps us (or any other plant or animal) from slumping into a pile of cells? And what keeps cells organized into tissues, and tissues into organs?
Multicellular organisms are made up of much more than cells. The cells in most tissues and organs are connected to a complex meshwork of proteins and polysaccharides known as the extracellular matrix. The architecture and structural integrity of a particular tissue or organ depends on these attachments between cells and the extracellular matrix. In this section, we explore the means by which cells are connected to one another and to proteins in the extracellular environment.
Cells are attached to one another and to the extracellular matrix by cell-surface proteins called cell adhesion molecules. The regions in the plasma membrane where cells make contact with and adhere to other cells or the extracellular matrix are called cellular junctions. Together, cell adhesion molecules, cellular junctions, and the extracellular matrix keep us (and all multicellular organisms) intact. The importance of cell adhesion is reflected by the fact that between 5% and 10% of the genes in the human genome are involved in attaching cells to one another or to the extracellular matrix.
10.4.1 Cell adhesion molecules allow cells to attach to other cells and to the extracellular matrix.
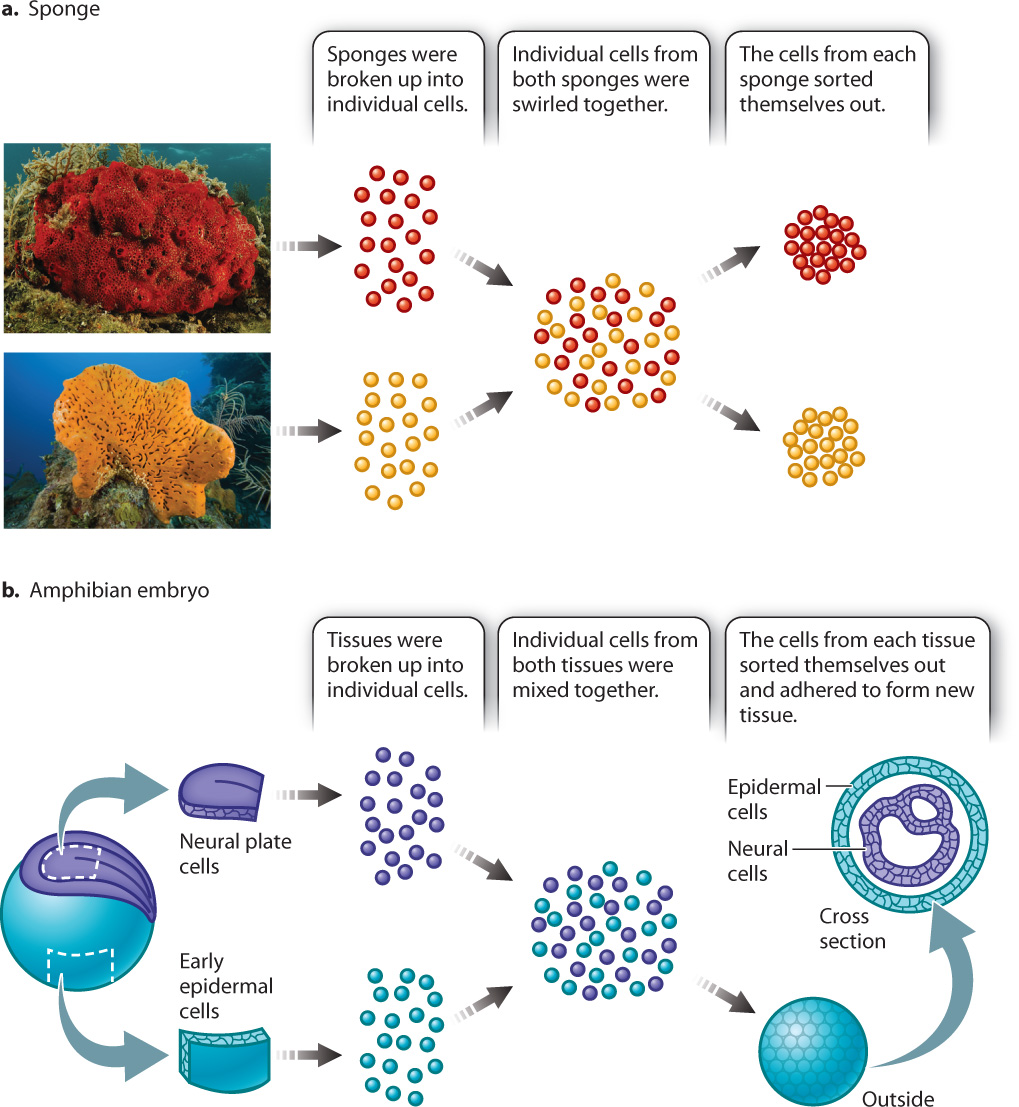
In 1907, American embryologist H. V. Wilson discovered that if he pressed a live sponge through fine cloth he could break up the sponge into individual cells. Then if he swirled the cells together, they would coalesce back into a group resembling a sponge. If he swirled together the cells from sponges of two different species, he observed that the cells sorted themselves out—that is, cells from one species of sponge associated only with cells from that same species (Fig. 10.15a). Fifty years later, German-born embryologist Johannes Holtfreter observed that if he took neuronal cells and skin cells from an amphibian embryo and treated them the same way that Wilson had treated sponge cells, the embryonic cells would sort themselves according to tissue type (Fig. 10.15b).
Cells can sort themselves because of the presence of various cell adhesion molecules. While a number of cell adhesion molecules are now known, the cadherins (for calcium-dependent adherence proteins) are especially important in the adhesion of cells to other cells. There are many different kinds of cadherins, and a given cadherin may bind only to another cadherin of the same type. This property explains Holtfreter’s observations of the cells from amphibian embryos. E-cadherin (for “epidermal cadherin”) is present on the surface of embryonic epidermal cells, and N-cadherin (for “neural cadherin”) is present on neuronal cells. The epidermal cells adhered to one another through E-cadherin, and the neuronal cells adhered to each other through N-cadherin.
Cadherins are transmembrane proteins (Chapter 5). The extracellular domain of a cadherin molecule binds to the extracellular domain of a cadherin of the same type on an adjacent cell (Fig. 10.16a). The cytoplasmic portion of the protein is linked to the internal cytoskeleton. This arrangement provides structural continuity from the cytoskeleton of one cell to the cytoskeleton of another, increasing the strength of tissues and organs.
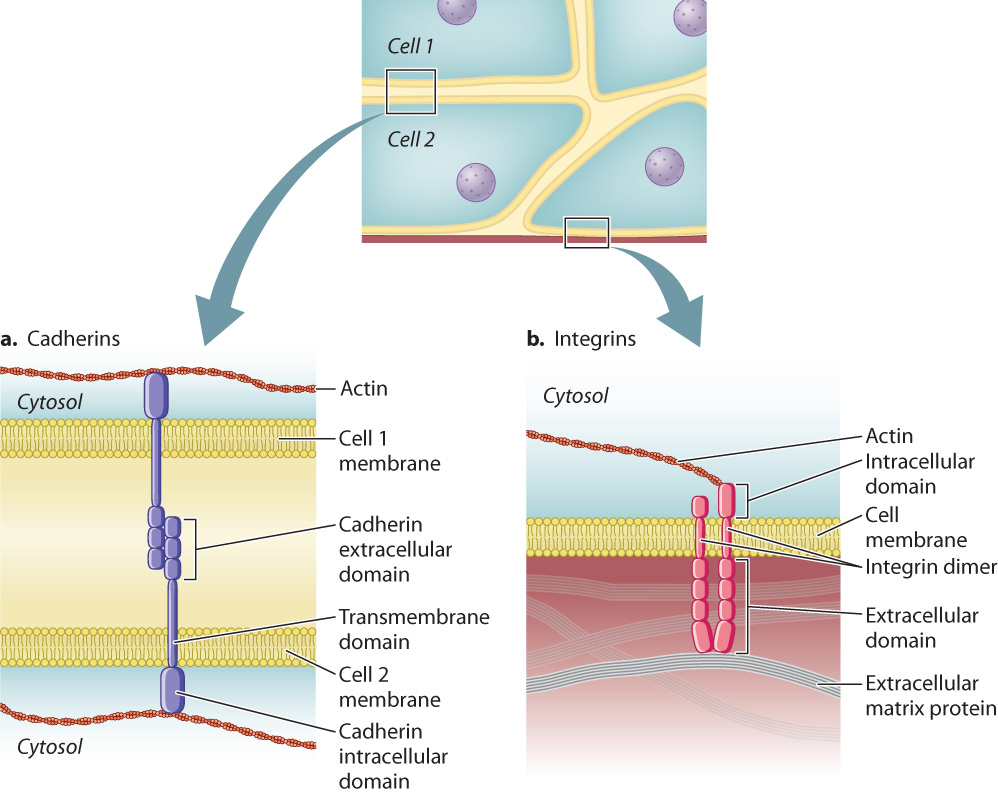
In addition to being stably connected to other cells, cells also attach to proteins of the extracellular matrix. This type of attachment provides structural reinforcement, especially to tissues under physical stress like the epidermis of the skin and the lining of the digestive tract. The cell adhesion molecules that enable cells to adhere to the extracellular matrix are called integrins. Like the cadherins, integrins are transmembrane proteins, and their cytoplasmic domain is linked to the cytoskeleton (Fig. 10.16b). Also like the cadherins, integrins are of many different types, each with a specificity for a different extracellular matrix protein. Integrins are present on the surface of virtually every animal cell. Though integrins have not been observed in plants, integrin-like proteins are present on the surface of some plant cells as well. We return to integrins and how they interact with the extracellular matrix in section 10.5.
10.4.2 Adherens junctions and desmosomes connect adjacent animal cells and are anchored to the cytoskeleton.
Cadherins are not distributed randomly in the plasma membrane but are located in adherens junctions and desmosomes. These junctional complexes anchor cells to one another and are reinforced by the cytoskeleton (Fig. 10.17).
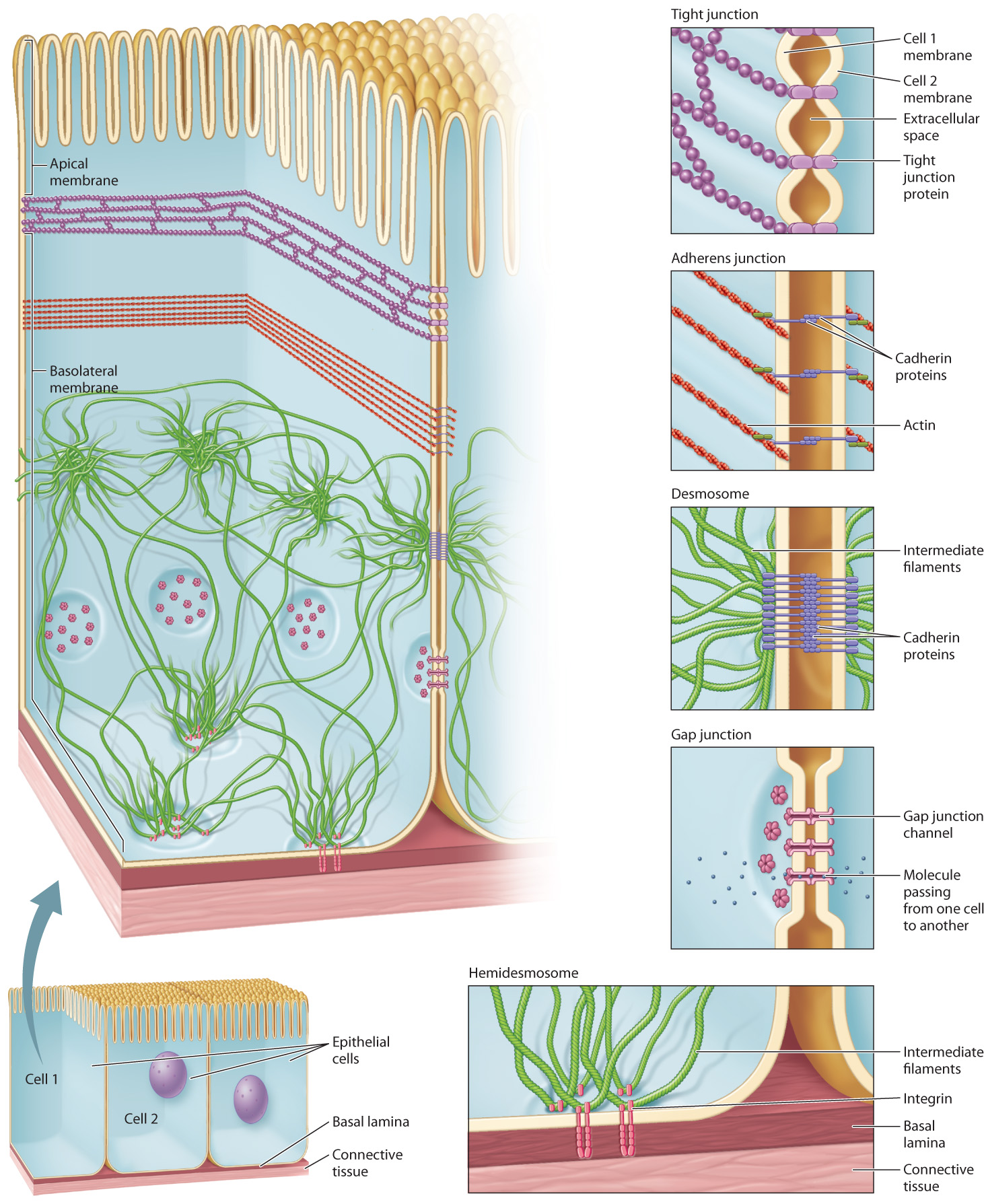
In our earlier discussion of microfilaments (section 10.2), we saw that a long bundle of actin microfilaments forms a band that extends around the circumference of epithelial cells, such as epithelial cells that line the intestine. This band of actin is attached to the plasma membrane by cadherins in a beltlike junctional complex called an adherens junction (Fig. 10.17). The cadherins in the adherens junction of one cell attach to the cadherins in the adherens junctions of adjacent cells. This arrangement establishes a physical connection among the actin cytoskeletons of all cells present in an epithelial layer of cells.
Desmosomes are buttonlike points of adhesion that hold the plasma membrane of adjacent cells together (Fig. 10.17). Cadherins are at work here, too, strengthening the connection between cells in a manner similar to adherens junctions. The cadherins in the desmosome of one cell bind to the cadherins in the desmosomes of adjacent cells. The cytoplasmic domain of these cadherins is linked to intermediate filaments in the cytoskeleton. This second type of physical connection among neighboring cells greatly enhances the structural integrity of epithelial cell layers.
The structural support provided by desmosomes and their associated network of intermediate filaments is crucial to the function of several organs, such as the skin and heart. Mutations in several of the genes for desmosomal proteins are responsible for a number of serious diseases. In these instances, intercellular connections are profoundly weakened. The results range from serious skin conditions due to fragile epidermis, as we saw with epidermolysis bullosa, to a high risk of early heart failure due to severe weakening of the heart muscle.
Quick Check 2
Adherens junctions and desmosomes both attach cells to other cells and are made up of cadherins. How, then, are they different?
10.4.3 Tight junctions prevent the movement of substances through the space between animal cells.
Epithelial cells, which form layers or sheets that cover or line other tissues and organs, define the boundaries of many of these tissues, including the digestive tract, respiratory tract, and outer layer of the skin. Like any effective boundary, a layer of epithelial cells must limit or control the passage of material across it. Adherens junctions and desmosomes provide strong connections between cells, but they do not prevent the free passage of materials through the spaces between the cells they connect. Junctional complexes called tight junctions (Fig. 10.17) establish a seal between cells so that the only way a substance can travel from one side of a sheet of epithelial cells to the other is by moving through the cells by means of one of the cellular transport mechanisms discussed in Chapter 5.
A tight junction is a band of interconnected strands of integral membrane proteins that, like adherens junctions, encircles the epithelial cell. It also binds to tight-junction proteins on adjacent cells. Unlike adherens junctions, however, tight junctions are not connected to the cytoskeleton. Their function is to prevent passage of materials in between cells, not to anchor the cells together.
Cells that have tight junctions have two distinct sides because the tight junction divides the plasma membrane into two distinct regions (Fig. 10.17). The portion of the plasma membrane in contact with the lumen, or the inside of any tubelike structure like the gut, is called the apical membrane. The apical membrane defines the “top” side of the cell. The rest of the plasma membrane is the basolateral membrane, which defines the bottom (“baso”) and sides (“lateral”) of the cell. These two regions of the plasma membrane are structurally different because the tight junction prevents lipids and proteins in the membrane on one side of the junction from diffusing to the other side. As a result, the apical and basolateral membranes of a cell are likely to have different integral membrane proteins, which causes them to be functionally different as well. In the small intestine, for example, glucose is transported from the lumen into intestinal epithelial cells by transport proteins on the apical side of the cells, and is transported out of the cells into the circulation by facilitated diffusion through glucose transporters restricted to the basolateral sides of the cells.
From this discussion, we see that cellular junctions and cytoskeletal elements interact to create stable communities of cells in the form of tissues and organs. These interactions are specific, with certain junctions associated with specific components of the cytoskeletal network. These interactions are summarized in Table 10.1.
| Table 10.1: Interactions among cellular junctions, cell adhesion molecules, and cytoskeleton elements | ||
|---|---|---|
| DIFFERENT TYPES OF CELLULAR JUNCTION | USE SPECIFIC CELL ADHESION MOLECULES | TO ANCHOR THEMSELVES TO THE CYTOSKELETON |
| Adherens junctions | Cadherins | Microfilaments |
| Desmosomes | Cadherins | Intermediate filaments |
| Hemidesmosomes | Integrins | Intermediate filaments |
10.4.4 Gap junctions and plasmodesmata allow the passage of substances from one cell to another.
Not all junctions are involved in anchoring cells in place or sealing off one side of a layer of cells from the other. The gap junctions (Fig. 10.17) of animal cells and plasmodesmata of plant cells are connections between the plasma membranes of adjacent cells that permit materials to pass directly from the cytoplasm of one cell to the cytoplasm of another. Gap junctions are formed when a set of integral membrane proteins arranged in a ring connects to a similar ring of proteins in the membrane of another cell. Ions and signaling molecules pass through these junctions, allowing cells to communicate. In the heart, for example, ions pass though gap junctions connecting cardiac muscle cells. This rapid electrical communication allows the muscle cells to beat in a well-timed, coordinated rhythm.
Plasmodesmata (the singular form is “plasmodesma”) are passages through the cell walls of adjacent plant cells. They are similar to gap junctions in that they allow cells to exchange ions and small molecules directly, but the similarity ends there. In plasmodesmata, the plasma membranes of the two connected cells are actually continuous. The size of the opening is considerably larger than that of gap junctions, large enough for cells to transfer RNA molecules and proteins, a capability that is especially important during plant embryonic development. Plasmodesmata allow plant cells to send signals to one another despite being enclosed within rigid cell walls.
Quick Check 3
Which type(s) of junction prevent substances from moving through the space between cells? Which one(s) permit the exchange of cytoplasmic materials?