11.3 MEIOTIC CELL DIVISION
As we have seen, mitotic cell division is important in the development of a multicellular organism and in the maintenance and repair of tissues and organs. Mitotic cell division is also the basis of asexual reproduction in unicellular eukaryotes. Sexual reproduction involves the fusion of gametes during fertilization to form a new organism. A new organism produced by sexual reproduction has the same number of chromosomes as its parents because the egg and sperm each have half the number of chromosomes as the diploid parents.
Gametes are produced by meiotic cell division, a form of cell division that includes two rounds of nuclear division. Meiotic cell division makes sexual reproduction possible. There are several major differences between meiotic cell division and mitotic cell division. First, meiotic cell division results in four daughter cells instead of two. Second, each of the four daughter cells contains half the number of chromosomes as the parent cell. (The word “meiosis” is from the Greek for “diminish” or “lessen.”) Third, the four daughter cells are each genetically unique. In other words, they are genetically different from each other and from the parental cell.
In multicellular animals, the cells produced by meiosis are the haploid eggs and sperm that fuse in sexual reproduction. In other organisms, such as fungi, the products are spores, and in some unicellular eukaryotes, the products are new organisms. In this section, we consider the steps by which meiosis occurs, its role in sexual reproduction, and how it likely evolved.
11.3.1 Pairing of homologous chromosomes is unique to meiosis.
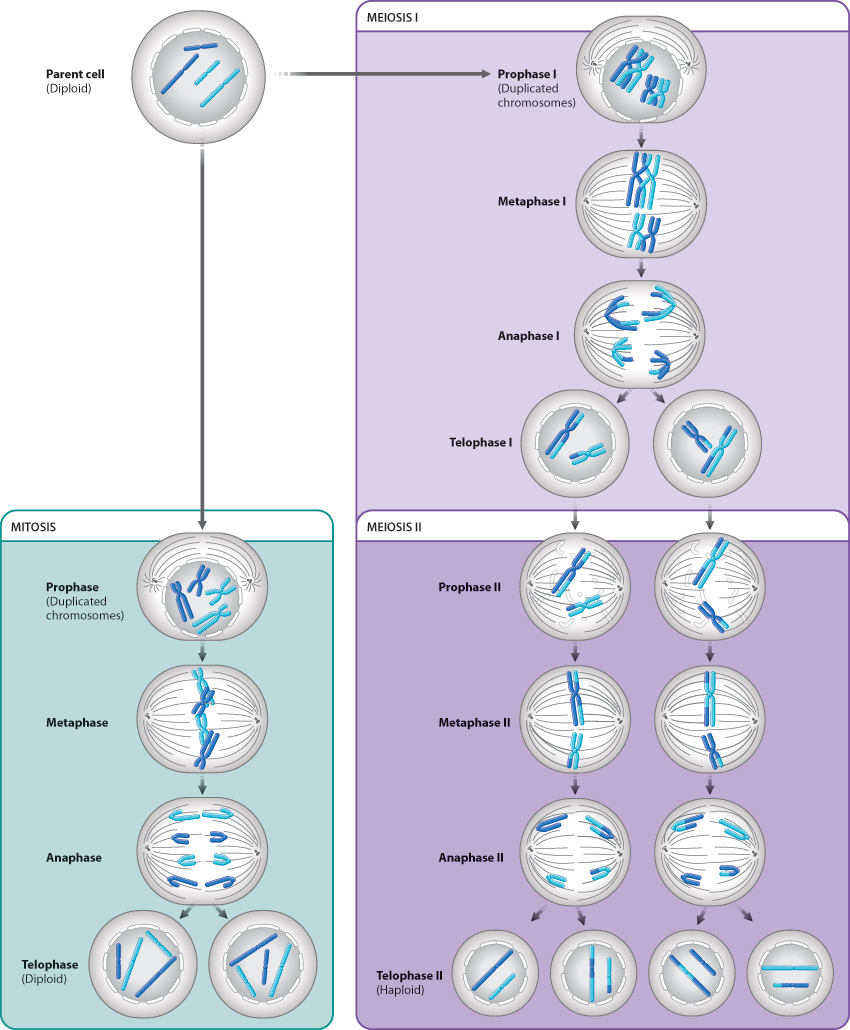
Like mitotic cell division, meiotic cell division follows one round of DNA synthesis, but, unlike mitotic cell division, meiotic cell division consists of two successive cell divisions. The two cell divisions are called meiosis I and meiosis II, and they occur one after the other. Each cell division results in two cells, so that by the end of meiotic cell division a single parent cell has produced four daughter cells.
Meiosis I begins with prophase I, illustrated in Fig. 11.8. The beginning of prophase I marks the earliest visible manifestation of chromosome condensation. The chromosomes first appear as long, thin threads present throughout the nucleus. By this time, DNA replication has already taken place, so each chromosome has become two sister chromatids held together at the centromere.
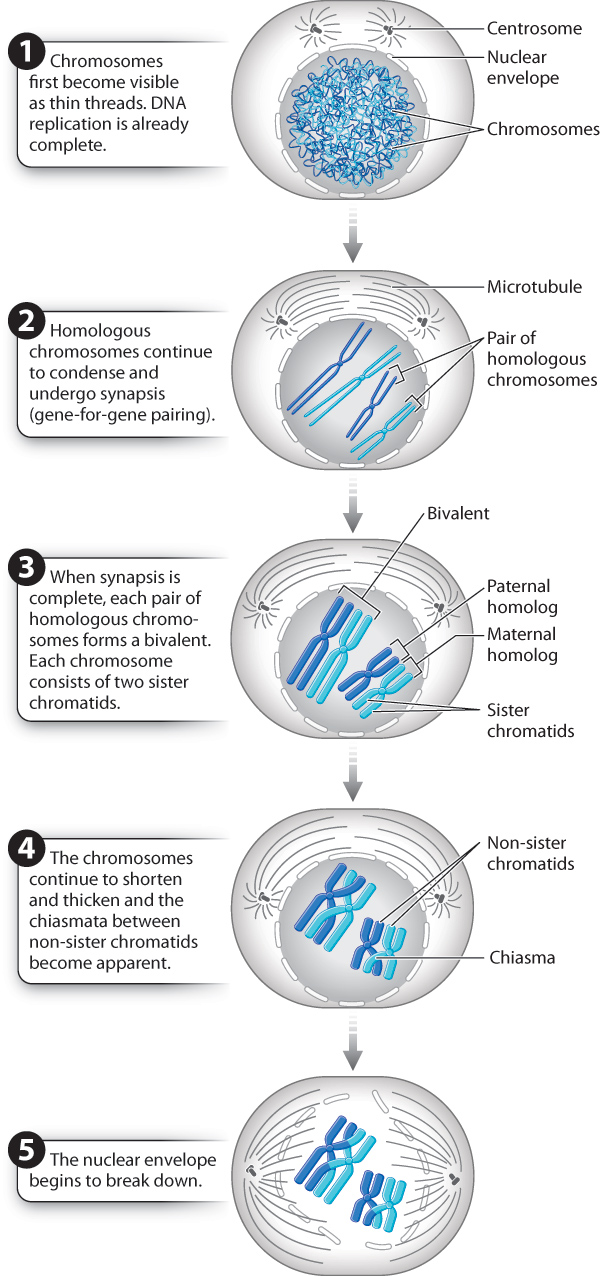
What happens next is an event of enormous importance and is unique to meiosis. The homologous chromosomes pair with each other, coming together to lie side by side, gene for gene, in a process known as synapsis. Even the X and Y chromosomes pair, but only at the tip where their DNA sequences are nearly identical. Because one of each pair of homologs is maternal in origin and the other is paternal in origin, chromosome pairing provides an opportunity for the maternal and paternal chromosomes to exchange genetic information, as described in the next section.
Because each homologous chromosome is a pair of sister chromatids attached to a single centromere, a pair of synapsed chromosomes creates a four-stranded structure: two pairs of sister chromatids aligned along their length. The whole unit is called a bivalent, and the chromatids attached to different centromeres are called non-sister chromatids, in contrast to sister chromatids, which result from replication of a single chromosome. The important distinction between sister chromatids and non-sister chromatids is that sister chromatids, as replicas of the same chromosome, are genetically identical; non-sister chromatids, which are replicas of different chromosomes, are genetically similar but not identical.
Quick Check 4
In a human cell at the end of prophase I, how many chromatids, centromeres, and bivalents are present?
11.3.2 Crossing over between DNA molecules results in exchange of genetic material.
Within the bivalents are crosslike structures, each called a chiasma (from the Greek meaning a “cross piece”; the plural is “chiasmata”) (Fig. 11.9). Each chiasma is a visible manifestation of a crossover, the physical breakage and reunion between non-sister chromatids.
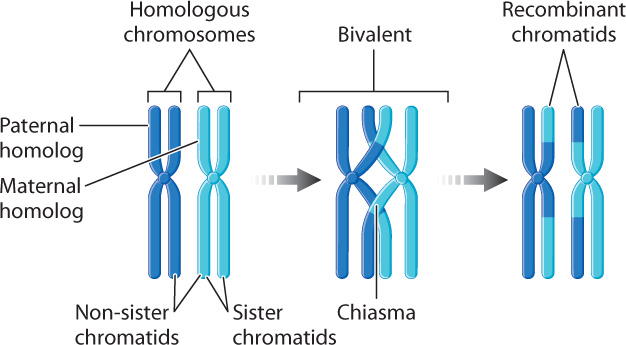
Through the process of crossing over, meiosis allows homologous chromosomes of maternal origin and paternal origin to undergo an exchange of DNA segments. The positions of these exchanges along the chromosome are essentially random, and therefore each chromosome that emerges from meiosis is unique, containing some DNA segments from the maternal chromosome and others from the paternal chromosome. The process is very precise: No nucleotides are gained or lost as homologous chromosomes exchange material. Note the results of crossing over as shown in Fig. 11.9: The recombinant chromatids are those that carry partly paternal and partly maternal segments. In this way, crossing over increases genetic diversity.
The number of chiasmata that are formed during meiosis depends on the species. In humans, the usual range is 50–60 chiasmata per meiosis. Most bivalents have at least one chiasma. Even the X and Y chromosomes are joined by a chiasma in the small region where they are paired. In addition to exchanging genetic material, the chiasmata also play a mechanical role in meiosis by holding the bivalents together while they become properly oriented in the center of the cell during metaphase, the stage we turn to next.
11.3.3 The first meiotic division brings about the reduction in chromosome number.
At the end of prophase I, the chromosomes are fully condensed and have formed chiasmata, the nuclear envelope has begun to disappear, and the meiotic spindle is forming. We are now ready to move through the remaining stages of meiosis I, which are illustrated in Fig. 11.10.
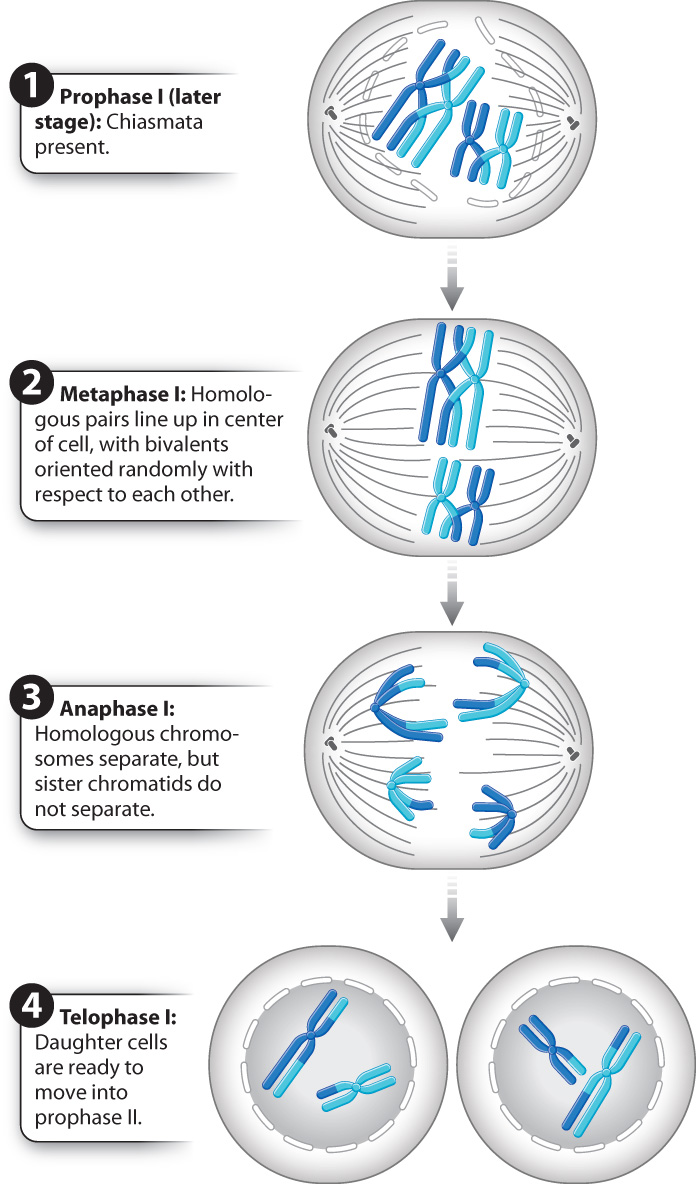
In metaphase I, the meiotic spindle is completed and the bivalents move so that they come to lie on an imaginary plane cutting transversely across the spindle. Each bivalent lines up so that its two centromeres lie on opposite sides of this plane, pointing toward opposite poles of the spindle. Importantly, the orientation of these bivalents is random with respect to each other. For some, the maternal homolog is attached to the spindle radiating from a given pole and the paternal homolog is attached to the spindle originating from the other pole. For others, the orientation is reversed. As a result, when the homologous chromosomes separate from each other in the next step (anaphase I), a complete set of chromosomes moves toward each pole, and that chromosome set is a random mix of maternal and paternal homologs. The random alignment of chromosomes on the spindle in metaphase I further increases genetic diversity in the products of meiosis.
At the beginning of anaphase I, the two homologous chromosomes of each bivalent separate as they are pulled in opposite directions. The key feature of anaphase I is that the centromeres do not split, and the two chromatids that make up each chromosome remain together. Anaphase I is thus very different from anaphase of mitosis, in which the centromeres split and each pair of chromatids becomes separated.
The end of anaphase I coincides with the arrival of the chromosomes at the poles of each spindle. Only one of the two homologous chromosomes goes to each pole, so in human cells there are 23 chromosomes at each pole at the end of meiosis I. Each of these chromosomes consists of two chromatids attached to a single centromere. Meiosis I is sometimes called the reductional division, since it reduces the number of chromosomes by half.
In telophase I, the chromosomes may uncoil slightly, a nuclear envelope briefly reappears, and in many species (including humans) the cytoplasm divides, producing two separate cells. The chromosomes do not completely decondense, however, and so telophase I blends into prophase II. Importantly, there is no DNA synthesis between the two meiotic divisions.
Quick Check 5
List three ways in which meiosis I differs from mitosis.
11.3.4 The second meiotic division resembles mitosis.
Now let’s turn to the second meiotic division, meiosis II, shown in Fig. 11.11. Starting with prophase II, the second meiotic division is in many respects like a conventional mitotic division, except that the nuclei in prophase II have the haploid number of chromosomes (23 in humans), not the diploid number. In prophase II, the chromosomes recondense to their maximum extent. Toward the end of prophase II, the nuclear envelope disappears (in those species in which it has formed), and the spindle begins to be set up.
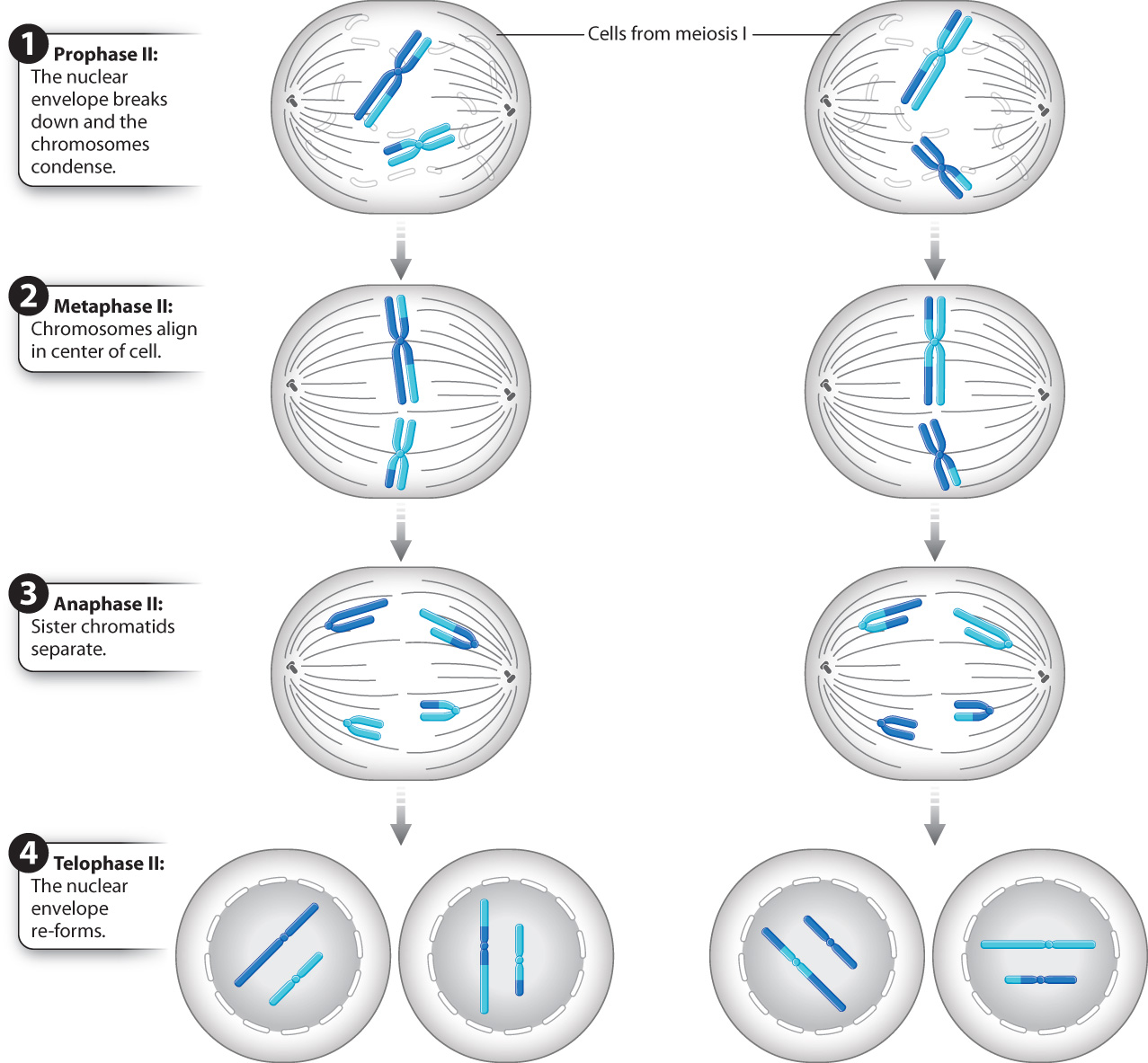
In metaphase II, the chromosomes line up so that their centromeres lie on an imaginary plane cutting across the spindle.
In anaphase II, the centromere of each chromosome splits. The separated chromatids, now each regarded as a full-fledged chromosome, are pulled toward opposite poles of the spindle. In this sense, anaphase II resembles anaphase of mitosis.
Finally, in telophase II, the chromosomes uncoil and become diffuse, a nuclear envelope re-forms around each set of chromosomes, and the cytoplasm divides by cytokinesis. Each resulting nucleus after telophase II has the haploid number of chromosomes. Because cells in meiosis II have the same number of chromosomes at the beginning and at the end, meiosis II is often called the equational division.
Quick Check 6
The genetic constitution of each cell after telophase II is different from the others. What two processes during meiosis result in these differences?
A comparison of mitosis and meiosis gives us hints about how meiosis might have evolved (Table 11.1 and Fig. 11.12). During meiosis I, maternal and paternal homologs separate from each other, whereas during meiosis II, sister chromatids separate from each other, similar to mitosis. The similarity of meiosis II and mitosis suggests that meiosis likely evolved from mitosis. Mitosis occurs in all eukaryotes and was certainly present in the common ancestor of all living eukaryotes. Meiosis is present in most, but not all, eukaryotes. Because the steps of meiosis are the same in all eukaryotes, meiosis is thought to have evolved in the common ancestor of all eukaryotes and has been subsequently lost in some lineages.
| Table 11.1: Comparison of Mitosis and Meiosis | ||
|---|---|---|
| MITOSIS | MEIOSIS | |
| Function | Asexual reproduction in unicellular eukaryotes Development in multicellular eukaryotes Tissue regeneration and repair in multicellular eukaryotes | Sexual reproduction Production of gametes and spores |
| Organisms | All eukaryotes | Most eukaryotes |
| Number of rounds of DNA synthesis | 1 | 1 |
| Number of cell divisions | 1 | 2 |
| Number of daughter cells | 2 | 4 |
| Chromosome complement of daughter cell compared to parent cell | Same | Half |
| Pairing of homologous chromosomes | No | Meiosis I—Yes Meiosis II—No |
| Crossing over | No | Meiosis I—Yes Meiosis II—No |
| Separation of homologous chromosomes | No | Meiosis I—Yes Meiosis II—No |
| Centromere division | Yes | Meiosis I—No Meiosis II—Yes |
| Separation of sister chromatids | Yes | Meiosis I—No Meiosis II—Yes |
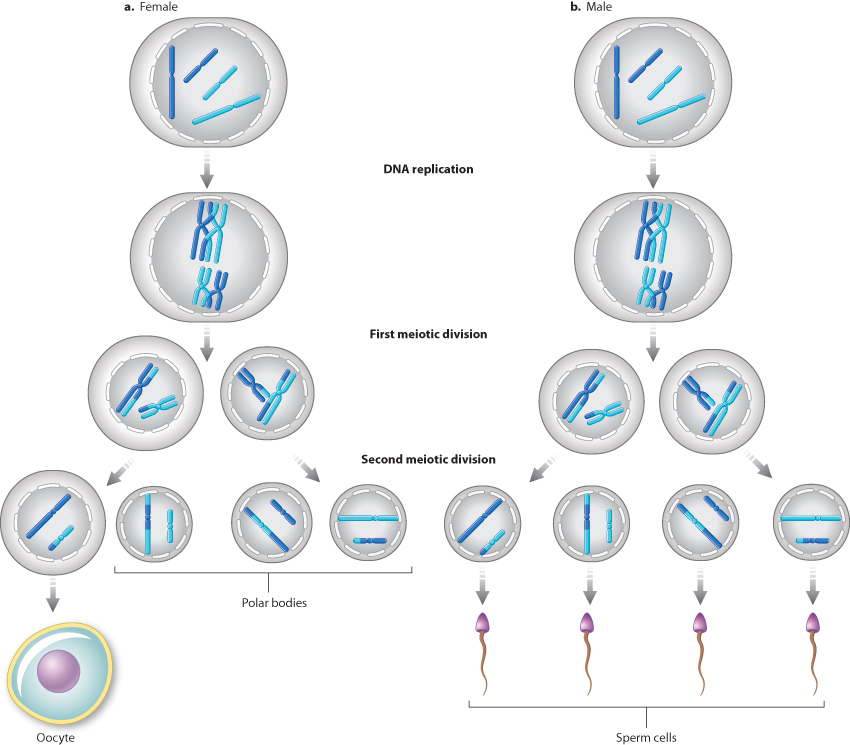
11.3.5 Division of the cytoplasm often differs between the sexes.
In multicellular organisms, division of the cytoplasm in meiotic cell division differs between the sexes. In female mammals (Fig. 11.13a), the cytoplasm is divided very unequally in both meiotic divisions. Most of the cytoplasm is retained in one meiotic product, a very large cell called the oocyte, which can develop into the functional egg cell, and the other meiotic products receive only small amounts of cytoplasm. These smaller cells are called polar bodies. In male mammals (Fig. 11.13b), the cytoplasm divides about equally in both meiotic divisions, and each of the resulting meiotic products goes on to form a functional sperm. During the development of the sperm, most of the cytoplasm is eliminated, and what is left is essentially a nucleus in the sperm head equipped with a long whiplike flagellum to help propel it toward the egg.
11.3.6 Meiosis is the basis of sexual reproduction.
Sexual reproduction involves two processes: meiotic cell division and fertilization. Meiotic cell division, as we just saw, produces cells with half the number of chromosomes present in the parent cell. In multicellular animals, the products of meiotic cell division are gametes: An egg cell is a gamete and a sperm cell is a gamete. Each gamete is haploid, containing a single set of chromosomes. In humans, meiosis takes place in the ovaries of the female and the testes of the male, and each resulting gamete contains 23 chromosomes, including one each of the 22 numbered chromosomes plus either an X or a Y chromosome.
During fertilization, these gametes fuse to form a single cell called a zygote. The zygote is diploid, having two complete sets of chromosomes, one from each parent. Therefore, fertilization restores the original chromosome number.
As we discuss further in Chapters 16 and 42, sexual reproduction plays a key role in increasing genetic diversity. Genetic diversity results from meiotic cell division (the cells that are produced are each genetically different from one another), and from fertilization (different gametes are combined to produce a new, unique individual). The increase in genetic diversity made possible by sexual reproduction allows organisms to evolve and adapt more quickly to their environment than is possible with asexual reproduction.
In the life cycle of multicellular animals like humans, then, the diploid organism produces single-celled gametes that fuse to make a diploid zygote. In this case, the only haploid cells in the life cycle are the gametes, and the products of meiotic cell division do not undergo mitotic cell division but instead fuse to become a diploid zygote. However, there are a number of life cycles in other organisms that differ in the timing of meiotic cell division and fertilization, discussed more fully in Chapter 27. Some organisms, like most fungi, are haploid. These haploid cells can fuse to produce a diploid zygote, but this cell immediately undergoes meiotic cell division to produce haploid cells, so that the only diploid cell in the life cycle is the zygote (Chapter 34). Other organisms, like plants, have both multicellular haploid and diploid phases (Chapter 30). In this case, meiotic cell division produces haploid spores that divide by mitotic cell division to produce a multicellular haploid phase, and subsequently haploid cells fuse to form a diploid zygote that also divides by mitotic cell division to produce a multicellular diploid phase.