Case 2 Cancer: When Good Cells Go Bad
11.5 What genes are involved in cancer?
As we have just seen, cells have evolved multiple regulatory mechanisms to ensure that they divide only at the appropriate time and place, and that cell division proceeds without error. What happens when cell division is not properly regulated? One possibility is that a cell divides rapidly, leading to cancer, a group of diseases characterized by uncontrolled cell division. When cells inappropriately activate mechanisms that promote cell division or lose the normal checks that prevent cell division, they can divide rapidly.
In this section, we explore the various ways in which cell cycle control can fail and lead to cancer. We begin with a discussion of a cancer caused by a virus. Although most cancers are not caused by viruses, the study of cancers caused by viruses helped us to understand how cancers develop.
11.5.1 Oncogenes promote cancer.
Our understanding of cancer is based partly on early observations of cancers in animals. In the first decade of the twentieth century, Peyton Rous studied cancers called sarcomas in chickens (Fig. 11.19). His work and that of others led to the discovery of the first virus known to cause cancer in animals, named the Rous sarcoma virus.
FIG. 11.19Can a virus cause cancer?
BACKGROUND In the early 1900s, little was known about the cause of cancer or the nature of viruses. Peyton Rous, an American pathologist, studied a form of cancer called a sarcoma in chickens. First, he moved a cancer tumor from a diseased chicken to a healthy chicken, and found that the cancer could be transplanted. Then, he tried to isolate the factor that causes the cancer in chickens. He made an extract of the tumor, filtered it to remove all the cells, and injected the extract into a healthy chicken to see if it could induce cancer.
HYPOTHESIS Experiments in other organisms, such as mice, rats, and dogs, showed that an extract free of cells from a tumor does not cause cancer. Therefore, Rous hypothesized that a cell-free extract of the chicken sarcoma would not induce cancer in healthy chickens.
EXPERIMENT Rous took a sample of the sarcoma from a chicken, ground it up, suspended it in solution, and centrifuged it to remove the debris. In one experiment, he injected this extract into a healthy chicken. In a second experiment, he passed the extract through a filter to remove all cells, including cancer cells and bacterial cells, and then injected this cell-free extract into a healthy chicken.
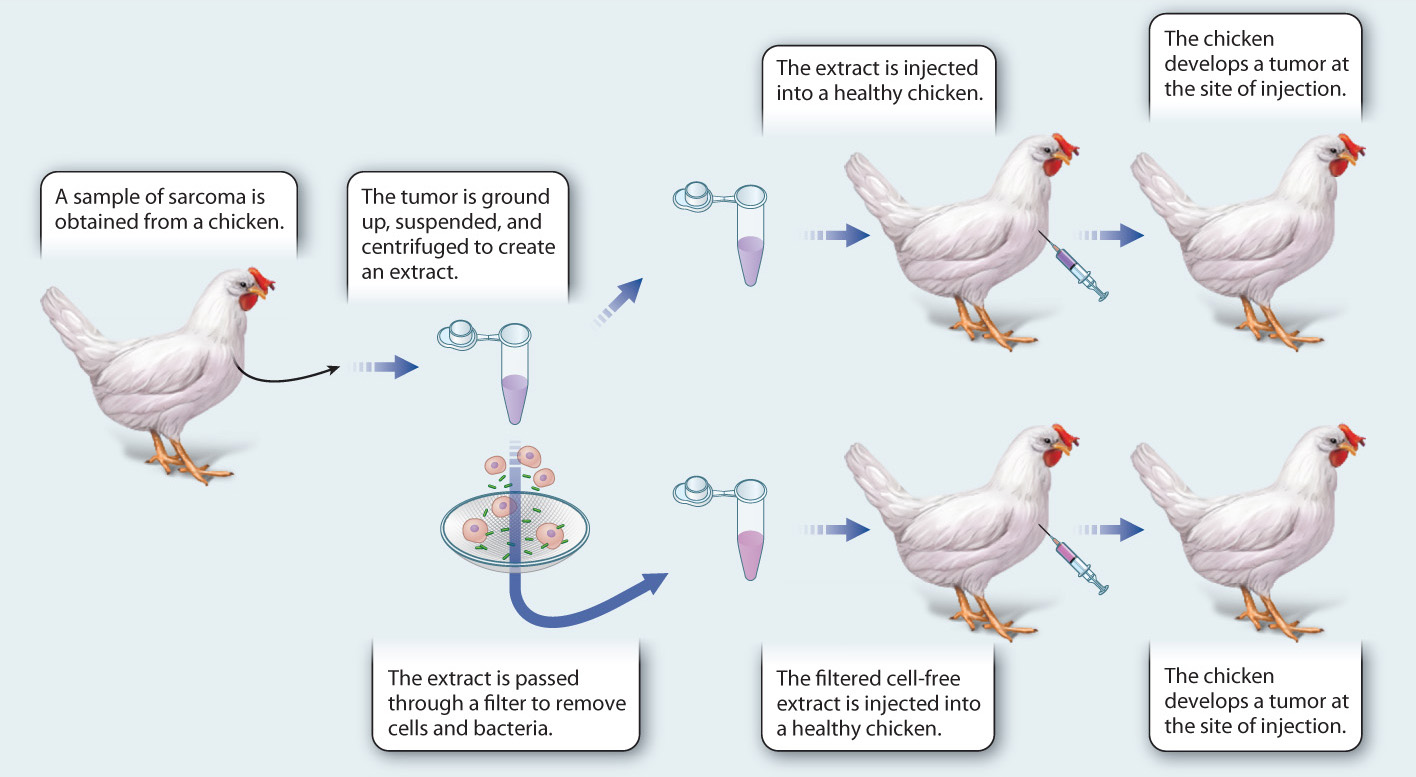
RESULTS In both cases, healthy chickens injected with the extract from the sarcoma developed cancer at the site of injection. Microscopic examination of the cancer showed it to be the same type of cancer as the original one.
CONCLUSIONS AND FOLLOW-UP WORK Rous concluded, contrary to his hypothesis, that a small agent—a virus or chemical—is capable of causing cancer. Later experiments confirmed that the cause of the cancer is a virus. This result was surprising, controversial, and dismissed at the time. A second cancer-causing virus was not found until the 1930s. Although most cancers are not caused by viruses, work with cancer-causing viruses helped to identify cellular genes that, when mutant, can lead to cancer. In 1966, Rous shared the Nobel Prize in Physiology or Medicine for his discovery.
SOURCES Rous, P. 1910. “A Transmissable Avian Neoplasm (Sarcoma of the Common Fowl).” J. Exp. Med. 12:696–705; Rous, P. 1911. “Transmission of a malignant new growth by means of a cell-free filtrate.” JAMA 56:198.
Viruses are assemblages of protein surrounding a core of either RNA or DNA. They multiply by infecting cells and using the biochemical machinery of their host to synthesize the proteins encoded in their genome and to make more copies of themselves. Viruses typically carry only a handful of genes, facilitating the identification of genes involved in cancer. Their investigation therefore provided a major breakthrough in our understanding of cancer.
As discussed in Chapter 9, growth factors activate several types of signaling proteins inside the cell that promote cell division. The gene from the Rous sarcoma virus that promotes uncontrolled cell division encodes a protein kinase similar to the cellular kinases that function as signaling proteins. This viral gene was named v-src, for viral-src (pronounced “sarc” and short for “sarcoma,” the type of cancer it causes).
Protein kinases are enzymes that activate other proteins by adding phosphate groups. These activation events are usually counterbalanced by other enzymes, called phosphatases, that remove phosphate groups. The viral protein encoded by v-src is overactive compared to cellular protein kinases. When the v-src gene is introduced into a host cell, it drives the cell to divide uncontrollably.
The v-src gene is one of several examples of an oncogene, or cancer-causing gene, found in viruses. A real surprise was the discovery that the v-src oncogene is found not just in the Rous sarcoma virus. It is in fact an altered version of a gene normally found in the host animal cell, known as c-src (cellular-src). The c-src gene plays a role during embryonic development and is involved in the control of cell division.
11.5.2 Proto-oncogenes are genes that when mutated may cause cancer.
The realization that the v-src oncogene has a normal counterpart in the host cell was an important step toward determining the cellular genes that participate in cell growth and division. These normal cellular genes are called proto-oncogenes because they are related to viral oncogenes but do not themselves cause cancer. Proto-oncogenes are normal genes important in cell division; only when they are mutated do they have the potential to cause cancer. Today, we know of scores of proto-oncogenes, most of which were identified through the study of cancer-causing viruses in chickens, mice, and cats.
Oncogenes also play a major role in human cancers. Most human cancers are not caused by viruses. Instead, human proto-oncogenes can be mutated into cancer-causing oncogenes by environmental agents such as chemical pollutants. A well-studied environmental factor linked to cancer is the set of aromatic amines present in the smoke of a burning cigarette. Aromatic amines are organic chemicals that can enter cells and associate with DNA, leading to damage or errors during DNA replication. Such changes result in mutations in DNA that can in some cases convert a normal proto-oncogene into an oncogene.
Not all proto-oncogenes encode protein kinases like Src. Nearly every protein that performs a key step in a signaling cascade that promotes cell division can be the product of a proto-oncogene. In other words, normal proto-oncogenes encoding growth factors, cell-surface receptors, G proteins, and protein kinases can be mutated to become oncogenes. Since the outcome of cell division depends on a complex and interconnected cascade of protein activities, any player in this scheme that does not function properly can lead to uncontrolled cell division.
Let’s consider an example of a proto-oncogene that codes for a cell-surface receptor. In Chapter 9, we discussed platelet-derived growth factor, or PDGF, which promotes cell division by binding to and dimerizing a receptor kinase in the membrane of the target cell. This action leads to the downstream activation of several signaling pathways, including the MAP kinase pathway and the promotion of cell division. In one type of leukemia (a cancer of blood cells), a mutation in the PDGF receptor gene results in an altered receptor that is missing the extracellular portion needed to bind the growth factor. Instead, the receptor dimerizes on its own, independent of PDGF binding. As a consequence, the receptor is always turned on.
Just like an overactive Src kinase, the overactive PDGF receptor turns on too many target proteins over too long a time period, leading to uncontrolled proliferation of blood cells. In both cases, mutations that change proto-oncogenes to oncogenes activate the protein inappropriately. That is, the mutant oncogene has more activity than its proto-oncogene counterpart.
11.5.3 Tumor suppressors block specific steps in the development of cancer.
From our discussion so far, you might think that oncogenes are solely responsible for the rampant cell growth that leads to cancer. This is true in some cases such as v-src in chickens, but more often something else must also go wrong before cancer can occur. Cells have safeguards that must be overcome before the controls on cell division are totally removed. These safeguards are a family of genes collectively called tumor suppressors because they encode proteins whose normal activities inhibit cell division.
Tumor suppressors prevent cell division by acting in opposition to proto-oncogenes. Therefore, whether a cell divides or not depends on the activities of both proto-oncogenes and tumor suppressors: Proto-oncogenes must be turned on and tumor suppressors must be turned off. Given the importance of controlling cell division, it is not surprising that cells have two counterbalancing systems that must be in agreement before division takes place.
We have already introduced a well-known example of a tumor suppressor, namely p53, which is defective in a majority of human cancers. As discussed earlier in this chapter and in Case 2: Cancer, the p53 protein normally halts cell division in response to DNA damage. When p53 activity is missing from the cell, excessive cell division is promoted.
Quick Check 8
How do oncogenes differ from tumor suppressor genes?
11.5.4 Most cancers require the accumulation of multiple mutations.
Most human cancers require more than the overactivation of one oncogene or the inactivation of a single tumor suppressor. Given the multitude of different tumor suppressor proteins that are produced in the cell, it is likely that one will compensate for even the complete loss of another. Cells have evolved a redundant arrangement of these control mechanisms to ensure that cell division is properly regulated.
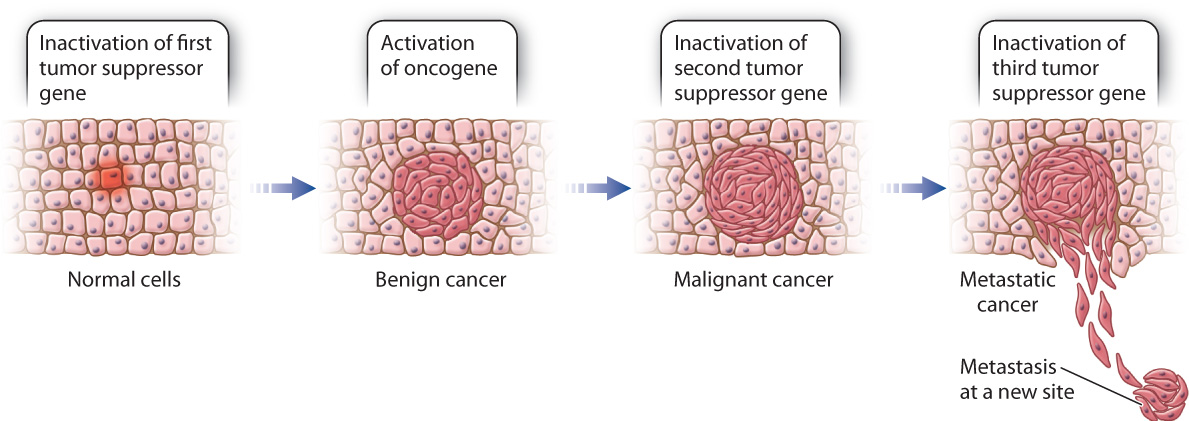
When several different cell cycle regulators fail, leading to both the overactivation of oncogenes and the loss of tumor suppressor activity, it is likely that cancer will develop. The cancer may be benign, which means that it is relatively slow growing and does not invade the surrounding tissue, or it may be malignant, which means that it grows rapidly and invades surrounding tissues. In many cases of malignant colon cancer, for example, tumor cells contain at least one overactive oncogene and several inactive tumor suppressor genes (Fig. 11.20). The gradual accumulation of these mutations over a period of years can be correlated with the stepwise progression of the cancer from a benign form to full malignancy.
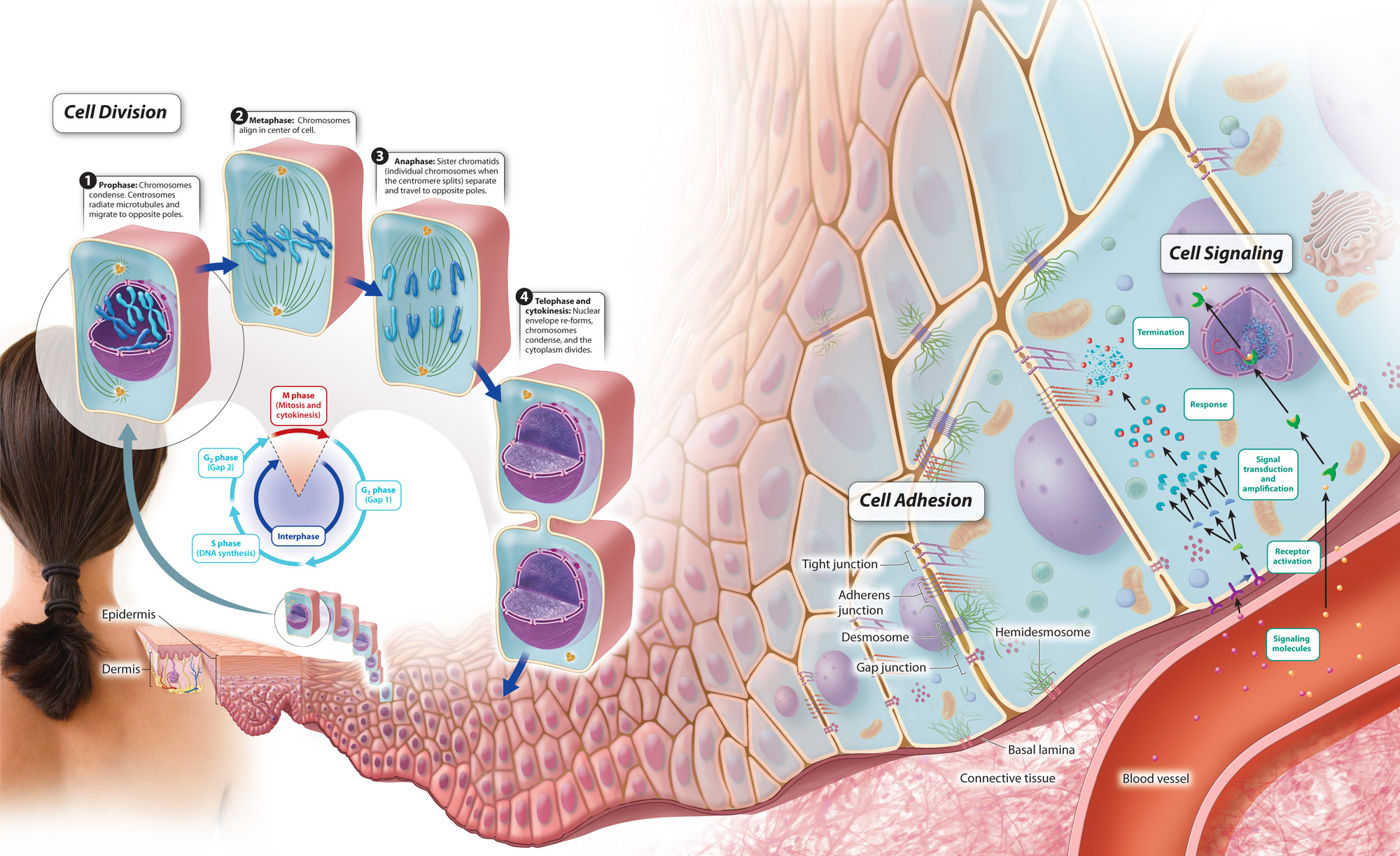
Taken together, we can now define some of the key characteristics that make a cell cancerous. Uncontrolled cell division is certainly important, but given the communities of cells and extracellular matrix we have discussed over the last few chapters, additional characteristics should also be considered. In 2000, American biologists Douglas Hanahan and Robert Weinberg wrote a paper called “Hallmarks of Cancer,” in which they highlighted key features of cancer cells. These include the ability to divide on their own in the absence of growth signals; resistance to signals that inhibit cell division and cell death; the ability to invade local and distant tissues (metastasis); and the production of signals to promote new blood vessel growth for nutrients to support cell division. Considered in this light, a cancer cell is one that no longer plays by the rules of a normal cellular community. Cancer therefore serves to remind us of the normal controls and processes that are required to allow cells to exist in a community. These processes, which work together, are summarized in Fig. 11.21 below.
Visual Synthesis: Cellular Communities
Click the image below to view an enlarged version in a new window.