14.4 DNA DAMAGE AND REPAIR
DNA is a fragile molecule, prone to damage of many different kinds. Every day, in each cell in our bodies, the DNA is damaged in some way at tens of thousands of places along the molecule. Fortunately, cells have evolved mechanisms for repairing various types of damage and restoring the DNA to its original condition. In this section, we examine some of the major types of DNA damage and some of the ways in which it is repaired.
14.4.1 DNA damage can affect both DNA backbone and bases.
Most mutations are spontaneous, occurring without an assignable cause. However, mutations can also be induced by radiation or chemicals. Mutagens are agents that increase the probability of mutation. The presence of a mutagen can increase the probability of mutation by a factor of 100 or more.
Some of the most important types of DNA damage induced by mutagens are illustrated in Fig. 14.16, which shows a highly mutated DNA molecule. Some types of damage affect DNA structure. These include breaks in the sugar–phosphate backbone, one of the main mutagenic effects of X-rays. Breaks can occur in just one strand of the DNA or both. Ultraviolet light can cause cross-links between adjacent pyrimidine bases, especially thymine, resulting in the formation of thymine dimers.
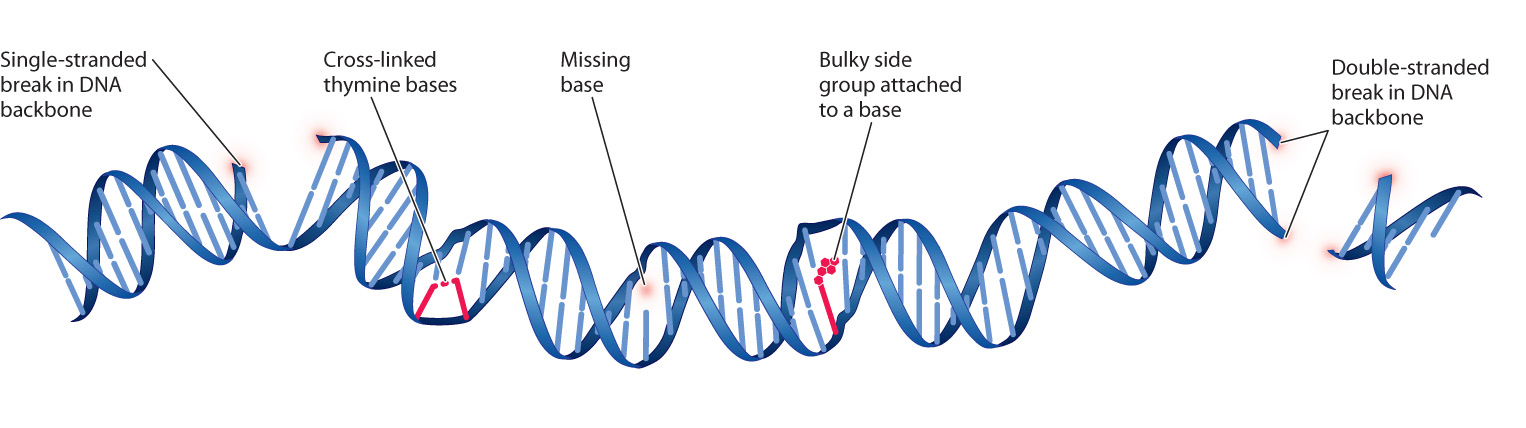
Yet another type of structural damage is loss of a base from one of the deoxyribose sugars, resulting in a gap in one strand where no base is present. Spontaneous loss of a purine base is one of the most common types of DNA damage, occurring at the rate of about 13,000 purines lost per human cell per day. The rate increases with age, and it can also be increased by exposure to oxidizing agents such as household bleach or hydrogen peroxide.
Other types of damage affect the bases themselves. Bases that are chemically damaged tend to mispair. Some bases are damaged spontaneously when reaction with a water molecule replaces an amino group (–NH2) with an atom of oxygen ( O). Some naturally occurring molecules mimic bases and can be incorporated into DNA and cause nucleotide substitutions. Caffeine mimics a purine base, for example (although to be mutagenic, the amount of caffeine required is far more than any normal person could possibly consume).
O). Some naturally occurring molecules mimic bases and can be incorporated into DNA and cause nucleotide substitutions. Caffeine mimics a purine base, for example (although to be mutagenic, the amount of caffeine required is far more than any normal person could possibly consume).
Chemicals that are highly reactive tend to be mutagenic, often because they add bulky side groups to the bases that hinder proper base pairing. The main environmental source of such chemicals is tobacco smoke. Other chemicals can perturb the DNA replication complex and cause the insertion or deletion of one or occasionally several nucleotides.
Quick Check 3
Earlier, we described the Lederbergs’ experiment, which demonstrated that mutations are not directed by the environment. But mutagens, which are environmental, can lead to mutations. What’s the difference?
14.4.2 Most DNA damage is corrected by specialized repair enzymes.
Cells contain many specialized DNA-repair enzymes that correct specific kinds of damage, and in this section we examine a few examples. Perhaps the simplest is the repair of breaks in the sugar–phosphate backbone, which are sealed by DNA ligase, an enzyme that can repair the break by using the energy in ATP to join the 3 hydroxyl of one end to the 5 phosphate of the other end. Most organisms have multiple different types of DNA ligase, some of which participate in DNA replication (Chapter 12) and others in DNA repair. One type of ligase seals single-stranded breaks in DNA, and a different type seals double-stranded breaks. Double-stranded breaks often result in point mutations or chromosomal rearrangements because they are less likely to be repaired than single-stranded breaks. In addition to their importance in DNA replication and repair, ligases are an important tool in research in molecular biology because they allow DNA molecules from different sources to be joined to produce recombinant DNA (Chapter 12).
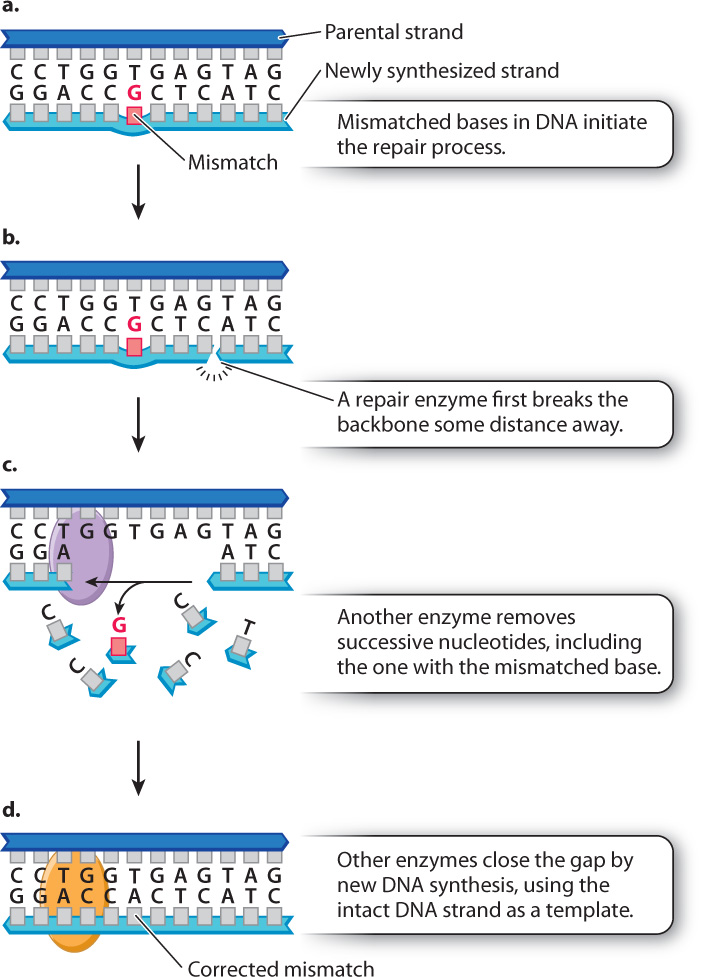
As we saw earlier in this chapter, mispairing of bases during DNA replication leads to the incorporation of incorrect nucleotides and potentially to nucleotide substitutions. Although 99% of the mispaired bases are corrected immediately by the proofreading function of DNA polymerase, another mechanism also scansthe DNA for mismatches to afford a second level of quality control. This process is called mismatch repair (Fig. 14.17). In mismatch repair, one of the strand backbones in the vicinity of the mismatch is cleaved and then degraded nucleotide by nucleotide to a point at a distance past the site of the mismatch. Then the gap is filled by new synthesis, which corrects the mismatch.
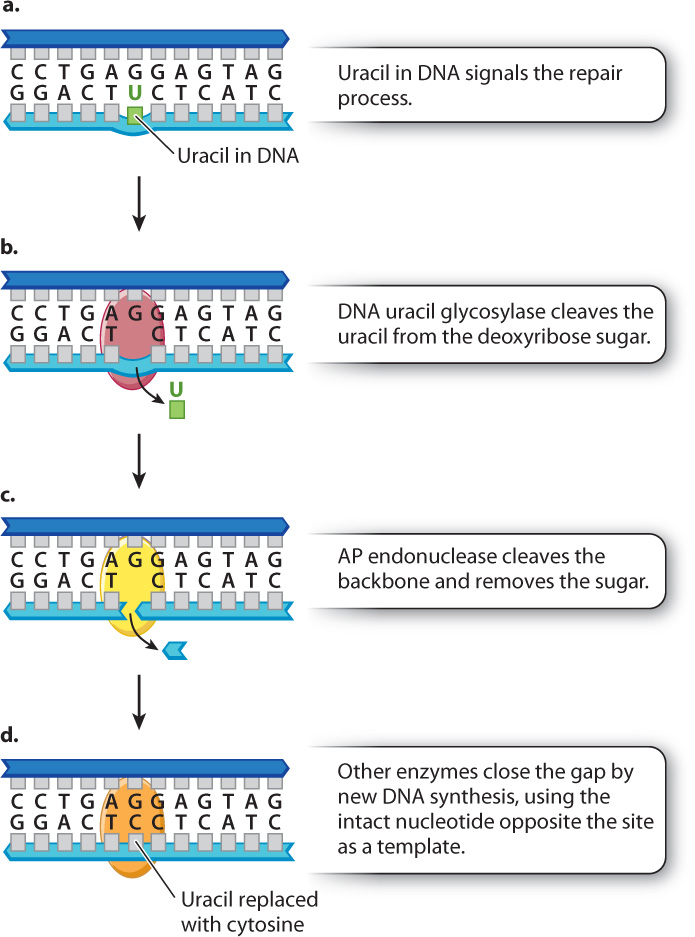
Fig. 14.18 shows an example of a more specialized repair system called base excision repair. Fig. 14.18a depicts a molecule in which uracil was mistakenly incorporated into a new DNA strand instead of cytosine. Recall that uracil is normally found in mRNA, but not in DNA. Most organisms have an enzyme called DNA uracil glycosylase that cleaves the uracil from its deoxyribose sugar, resulting in a sugar in the DNA backbone lacking a base (Fig. 14.18b). Such sites are recognized by an enzyme called AP endonuclease (Fig. 14.18c), which cleaves the backbone on both sides of the sugar that lacks a base. This cleavage results in a one-nucleotide gap, which is filled by new synthesis using the intact DNA strand as a template. The result is that uracil is removed and replaced with cytosine. Fig. 14.18 shows DNA repair systems that work in tandem, in this case DNA uracyl glycosylase acting with the AP endonuclease. Other types of glycosylases remove abnormal or damaged bases from the DNA, and these enzymes are also coupled with the AP endonuclease.
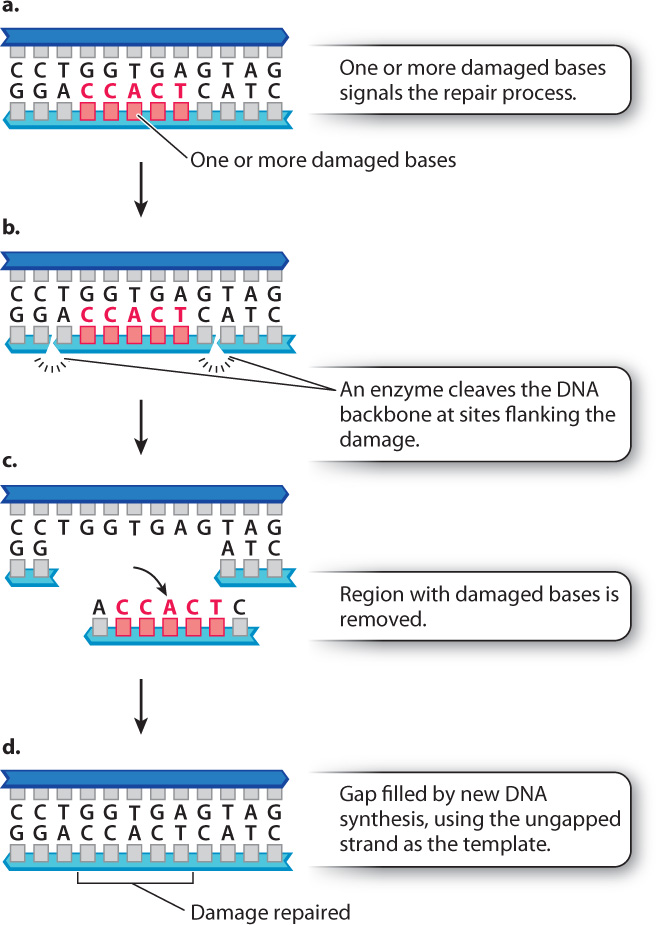
Nucleotide excision repair, which is similar in some ways to mismatch repair, is illustrated in Fig. 14.19. The major difference is that mismatch repair operates on single mispaired bases, whereas nucleotide excision repair recognizes multiple mismatched or damaged bases across a region. The region excised in mismatch repair is small, but that in nucleotide excision repair can extend for thousands of nucleotides. Nucleotide excision repair is also used to remove nucleotides with some types of bulky side group, as well as thymine dimers resulting from ultraviolet light.
The importance of DNA repair mechanisms is illustrated by considering what happens when they do not function properly. Xeroderma pigmentosum (XP) is a disease in which nucleotide excision repair is defective. People with XP are exquisitely sensitive to the UV radiation in sunlight. Because of the defect in nucleotide excision repair, damage to DNA resulting from UV light is not corrected, leading to the accumulation of mutations in skin cells. The result is high rates of skin cancer. People with XP must minimize their exposure to sunlight and in extreme cases stay out of the sun altogether.
When DNA repair mechanisms function properly, they reduce the rate of mutation to a level that is compatible with life. But they are not perfect, and they do not catch all the mistakes. The result is genetic variation, the raw material of evolution.