24.3 DISTINCT FEATURES OF OUR SPECIES
Many extraordinary changes in anatomy and behavior occurred in the 5–7 million years since our lineage split from the lineage that gave rise to the chimpanzees. Fossils tell us a great deal about those changes, especially when high-quality material such as Lucy or Ardi is available, but in general this is an area in which fossils are hard to come by and there is a lot of speculation. Speculation is especially common when we try to explain the reasons behind the evolution of a particular trait. Why, for example, did language evolve? It is easy enough to think of a scenario in which natural selection favors some ability to communicate—maybe language arose to facilitate group hunting. There are plenty of plausible ideas on the subject, but, in most cases, no evidence, so it is impossible to distinguish among competing hypotheses. We can, however, be confident that the events that produced language occurred in Africa, and, through paleontological studies of past environments, we can conclude that humans evolved in an environment similar in many ways to today’s East African savanna.
24.3.1 Bipedalism was a key innovation.
The shift from walking on four legs to walking on two was probably one of the first changes in our lineage. Many primates are partially bipedal—chimpanzees, for example, may be observed knuckle-walking—but ours is the only species that is wholly bipedal. As we’ve seen, Lucy, dating from about 3.2 million years ago, was already bipedal, and 4.4 million years ago, Ardi was partially bipedal. We can further refine our estimate of when full bipedalism arose from the evidence of a trace fossil. A set of 3.5-million-year-old fossil footprints discovered in Laetoli, Tanzania, reveal a truly upright posture.
Becoming bipedal is not simply a matter of standing up on hind legs. The change required substantial shifts in a number of basic anatomical characteristics, described in Fig. 24.11.
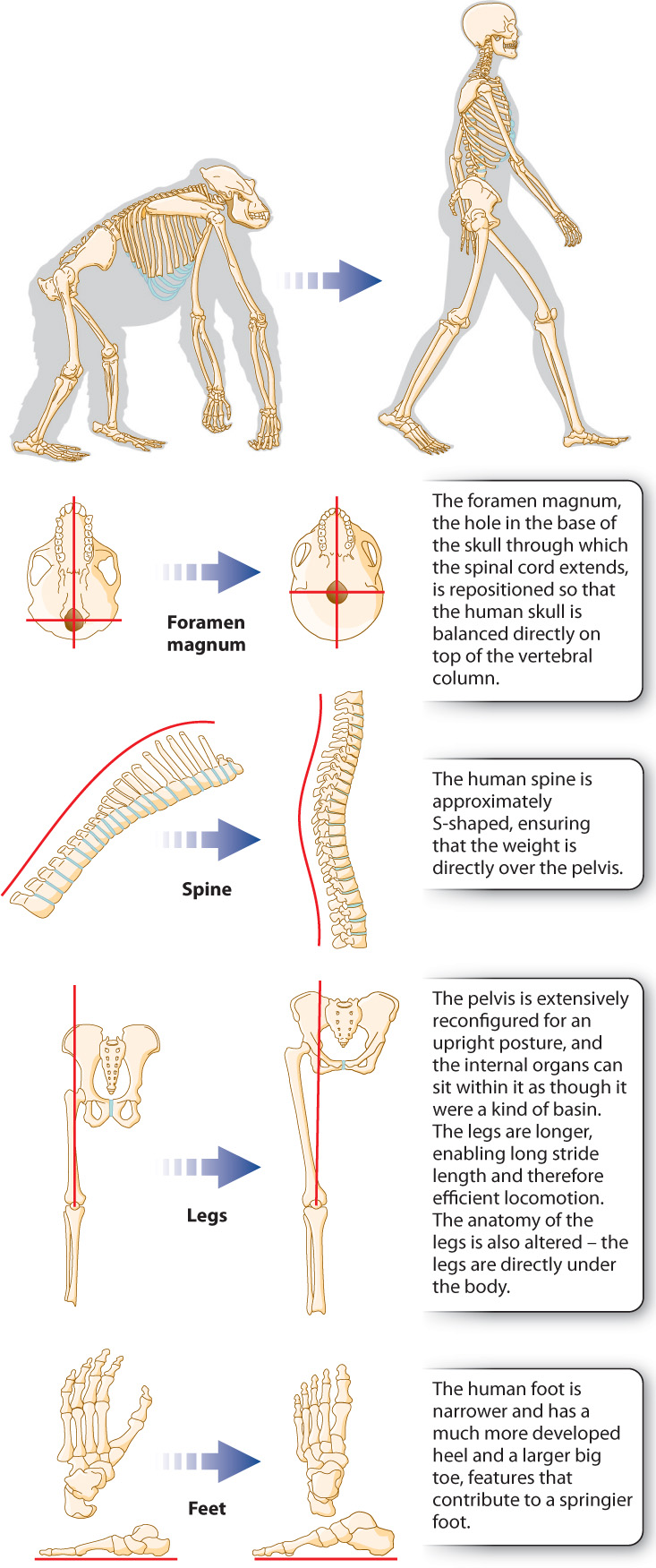
Why did hominins become bipedal? Maybe it gave them access to berries and nuts located high on a bush. Maybe it allowed them to scan the vicinity for predators. Maybe it made long-distance travel easier. And, very important, bipedalism freed up our ancestors’ hands. No longer did they need their hands for locomotion, so for the first time there arose the possibility that specialized hand function could evolve. Most primates have some kind of opposable thumb, but the human version is much more refined. The human thumb has three muscles that are not present in the thumb of chimpanzees, and these allow much finer motor control of the thumb. Tool use, present but crude in chimpanzees, can be much more subtle and sophisticated with a human hand.
Bipedalism also made it possible to carry material over long distances. Hominins could then set up complex foraging strategies whereby some individuals supply others with resources. Exactly when sophisticated tool use arose in our ancestors is controversial, but it is indisputable that bipedalism contributed. Similarly, the ability to manipulate food with the hands, and to carry material using the hands rather than the mouth, likely permitted the evolution of the human jaw—indeed, the entire facial structure—in such a way that language became a possibility.
24.3.2 Adult humans share many features with juvenile chimpanzees.
King and Wilson’s 1975 discovery that the DNA of humans and chimpanzees are 99% identical has recently been confirmed by DNA sequence comparison of the human and chimpanzee genomes. It turns out that the two genomes are extraordinarily similar: All our genes are also present in the chimpanzee and their sequences are extremely similar, suggesting that the functions of the proteins they code for are the same. If we are so similar, how can we account for the extensive differences between the two species?
In their original paper, King and Wilson suggested that much of the most significant evolution along the hominin lineage came about through changes in the regulation of genes (Chapter 19). A small change—one that causes a gene to be transcribed at a different stage of development, for example—can have a major effect. In other words, King and Wilson introduced a model whereby small changes in the software could have a major impact even though the basic hardware is the same.

One of the gene-regulatory pathways that changed may be responsible for human neoteny, the long-term evolutionary process in which the timing of development is altered so that a sexually mature organism still retains the physical characteristics of the juvenile form. As early as 1836, French naturalist Étienne Geoffroy Saint-Hilaire noted that a young orangutan on exhibit in Paris looked considerably more like a human than the adult of its own species (Fig. 24.12).
Several human attributes support this model, including our large heads (and correspondingly large brains), a feature of juvenile great apes. A second human attribute is our lack of hair. The juveniles of other great apes are not as hairless as humans, but they’re considerably less hairy than adult apes. A third attribute is the position of the foramen magnum at the base of the skull. In primate development, the foramen magnum starts off in the position it occupies in the adult human and then, in nonhuman great apes, migrates toward the back of the skull. Adult humans have retained the juvenile great ape foramen magnum position. Finally, it has been suggested that our mentality, with its questioning and playfulness, is equivalent in many ways to that of a juvenile ape rather than to that of the comparatively inflexible adult ape.
In keeping with King and Wilson’s idea, this shift in development could conceivably be achieved with relatively little genetic change. All that it might take would be a few changes to the regulatory switches that control the timing of development.
24.3.3 Humans have large brains relative to body size.
In mammals, brain size is typically correlated with body size. Humans are relatively large-bodied mammals, but our brains are large for even our body size (Fig. 24.13). It is our large brains that have allowed our species’ success, extraordinary technological achievements, and at times destructive dominion over the planet.
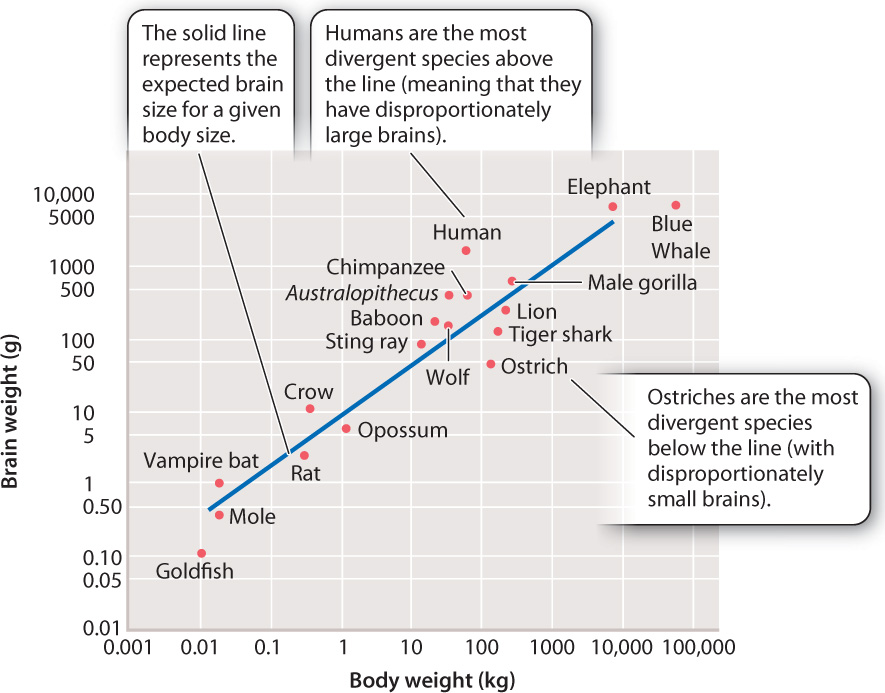
What factors promoted the evolution of the large human brain? Again, speculation is common, and it is unlikely we will ever have a definitive answer. Because a large brain is metabolically expensive to produce and to maintain, we can conclude that natural selection must have acted in favor of large brains. What are the selective factors? Here are some possibilities:
- Tool use. Bipedalism permitted the evolution of manual dexterity, which in turn requires a complex nervous organization if delicate hands are to be useful.
- Social living. Groups require coordination, and coordination requires some form of communication and the means of integrating and acting upon the information conveyed. One scenario, for example, sees group hunting as critical in the evolution of the brain: Natural selection favored those individuals who cooperated best as they pursued large prey.
- Language. Did the evolution of language drive the evolution of large brains? Or did language arise as a result of having large brains? Again, we will probably never know, but it is tempting to speculate that the brain and our extraordinary powers of communication evolved in concert.
Probably, as with bipedalism, there was no single factor but a mix of elements that worked together to result in the evolution of a large brain size. We tend to focus on brain size because it is a convenient stand-in for mental power and because we can measure it in the fossil record by making the reasonable assumption that the volume of a fossil’s cranium reflects the size of its brain. However, it should be emphasized that our minds are not the products solely of larger brains. Rather, the reorganization of existing structures and pathways is the more important product of the evolution of the brain. Not only was the brain expanding over those 7 million years of human evolution, it was also being rewired.
The human brain evolved through natural selection. What we have today is a learning machine capable of generating many more skills and abilities than just those that directly enhance evolutionary fitness. For example, let’s assume that the brain evolved to make group hunting more efficient. The result is that the brain does indeed enhance our ability to hunt together, but there are numerous by-products of this brain that take its abilities beyond just hunting. Playing the piano, for instance, has nothing to do with group hunting, and yet the brain allows us to do it. A brain evolved for a mundane task like group hunting lies at the heart of much that is wonderful about humanity.
24.3.4 The human and chimpanzee genomes help us identify genes that make us human.
The key genetic differences between humans and chimpanzees must lie in the approximately 1% of DNA sequence that differs between the two genomes. What do we see when we compare human and chimpanzee genomes?
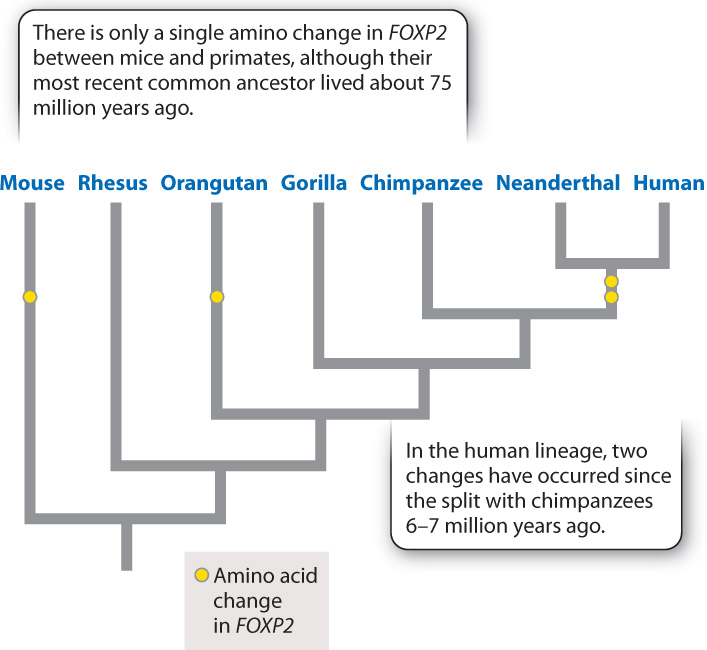
A gene that has attracted a lot of interest is FOXP2, a member of a large family of evolutionarily conserved genes that encode transcription factors that play important roles in development. Individuals with mutations in FOXP2 often have difficulty with speech and language. Interestingly, other animals whose FOXP2 has been knocked out also have communication impairments. Songbirds are less capable of learning new songs, and the ultrasonic “songs” of mice are disrupted. In addition to its effects on the brain, FOXP2 plays a role in the development of many tissues.
Studies of the gene’s amino acid sequence reveal a pattern of extreme conservation. The sequences in mice and chimpanzees, whose most recent common ancestor lived about 75 million years ago, differ by a single amino acid (Fig. 24.14). However, two amino acids are present in humans but absent in chimpanzees. Studies of Neanderthal DNA have shown that Neanderthals possessed the modern human version of FOXP2. The presence of the same version of the gene in both species suggests that the differences arose in the hominin line before the split between our species and Neanderthals, which occurred at least 600,000 years ago.
FOXP2 gives us a glimpse of the genetic architecture of the traits that are likely to be important in the determination of “humanness.” Those two differences in amino acid sequence are intriguing, but they do not a human make. Substitute the human FOXP2 gene into a mouse and you will not get a talking mouse. The critical genetic differences are many, subtle, and interacting.
Quick Check 4
The FOXP2 gene is sometimes called the “language gene.” Why is this name inaccurate?