27.4 THE FOSSIL RECORD OF PROTISTS
Not all algae and protozoa fossilize easily, but those that do provide a striking record of evolution that both predates and parallels the better-known fossil record of animals. Fossilized protists do not contain DNA, nor do they preserve the features of cell biology that define eukaryotes, such as nuclear membranes and organelles. Thus, for a fossil to be identified as eukaryotic, it must preserve other features of morphology found today in Eukarya, but not in Bacteria or Archaea. For this reason, fossils that can be identified with certainty as eukaryotic are limited to those left by eukaryotes that synthesized distinctive and preservable cell walls at some stage of their life cycle.
27.4.1 Fossils show that eukaryotes existed at least 1800 million years ago.

Sedimentary rocks deposited in coastal marine environments 1800–1400 million years ago contain microscopic fossils (called microfossils for short) up to about 300 μm in diameter. Many of these have complicated wall structures that identify them as eukaryotic. Interlocking plates, long and branching arms, and complex internal layering have been imaged by both scanning and transmission electron microscopes (Fig. 27.23a). Comparison with living organisms suggests that such fossils could only be formed by organisms with a cytoskeleton and endomembrane system, the hallmarks of eukaryotic biology.
At present, it appears that the eukaryotic domain first appeared no later than 1800 million years ago and possibly much earlier, although the record is difficult to trace in older rocks. None of these earliest eukaryotic microfossils can be placed with confidence into one of the living superkingdoms. Conceivably, at least some of the oldest protistan microfossils could predate the last common ancestor of living eukaryotes.
In sedimentary rocks thought to be about 1200 million years old, we find the oldest fossils that can be linked clearly to a living group of eukaryotes (Fig. 27.23b). These fossils preserve features found today only in the red algae. We can therefore conclude that by 1200 million years ago Archaeplastida had already diverged from other eukaryotic superkingdoms, photosynthesis had become established in eukaryotes, and simple multicellularity had already evolved within the domain.
Much more eukaryotic diversity is recorded in rocks 800–700 million years old, including green algae and the remains of test-forming amoebozoans (Fig. 27.23c). Molecular fossils further indicate the presence of ciliates and dinoflagellates in the emerging eukaryotic world and show that algae were expanding to become major photosynthesizers in the oceans. Until about 800 million years ago, subsurface waters of the oceans were commonly poorly supplied with oxygen and were sometimes rich in sulfide, as well. After this time, sulfide appears to have declined while oxygen increased. It may be, then, that eukaryotic organisms diversified as the oceans and atmosphere began to draw closer to their modern states.
27.4.2 Protists have continued to diversify during the age of animals.
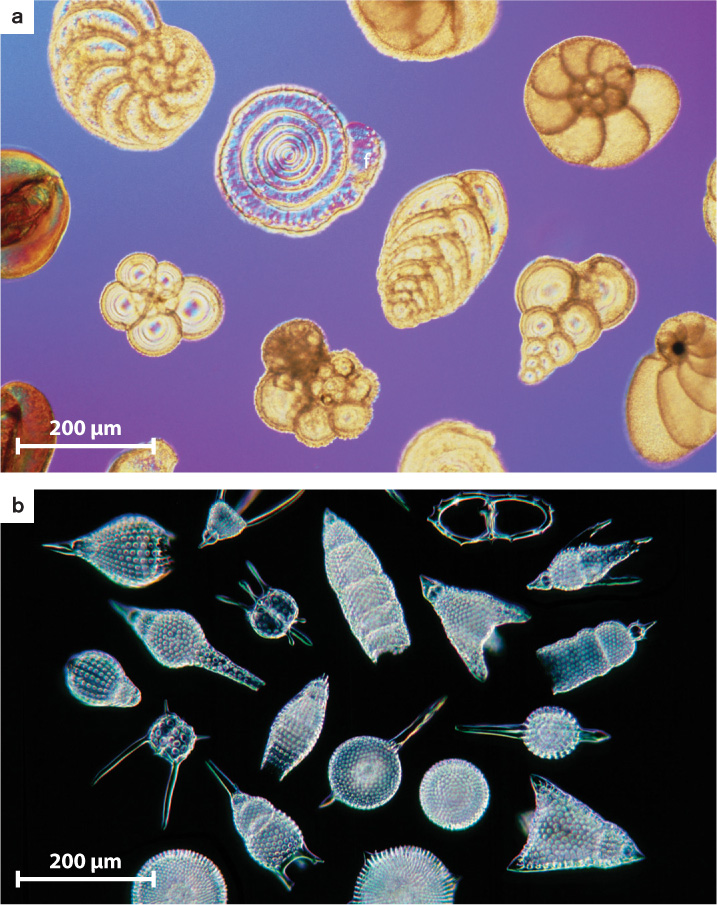
Modern protistan diversity took shape alongside evolving animals during the Phanerozoic Eon, 542 million years ago to the present. For example, foraminiferans and radiolarians, marine protozoans with shells of calcium carbonate and silica, respectively, appear for the first time in rocks deposited near the beginning of the Phanerozoic Eon and have continued to diversify since that time (Fig. 27.24). Some algal seaweeds also evolved skeletons of calcium carbonate at this time, which diversified along with animals in Phanerozoic oceans. Why did algae and protozoans evolve mineralized skeletons? Presumably to keep from being eaten.
Marine protists underwent a new round of radiation during the Mesozoic Era, 252–65 million years ago. The most significant changes occurred among photosynthesizers. During the preceding Paleozoic Era, green algae and cyanobacteria were the primary photosynthesizers in the oceans, but as the Mesozoic Era began, new groups of photosynthetic dinoflagellates and coccolithophorid algae appeared, followed by diatoms. Today, these three groups dominate photosynthesis in many parts of the sea.
Within the oceans, diatom and coccolithophorid evolution had another important consequence. The silica skeletons of diatoms are now the principal means by which SiO2 is transferred from seawater to sediments. Similarly, the CaCO3 scales made by coccolithophorids and the skeletons of planktonic foraminifera are a primary source of carbonate sediments on the seafloor (Fig. 27.25). In this way, the ongoing evolution of protists in the age of animals has continued to transform marine ecosystems.
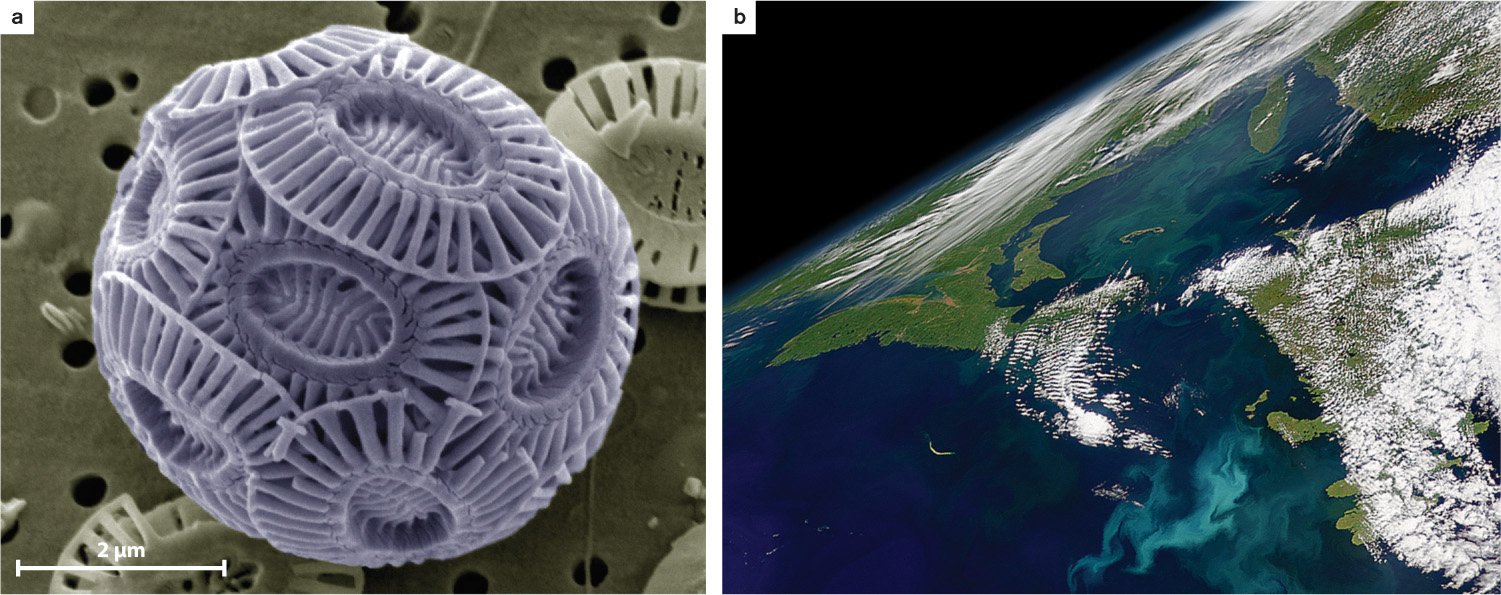
Today, microscopic eukaryotes dominate photosynthetic production in coastal oceans and play a major role in biogeochemical cycles. And for thousands of protists, plants and animals are simply habitats, ripe for colonization—we need look no further than the many eukaryotic vectors of human disease. The key conclusion is that a remarkable, and still largely undocumented, diversity of eukaryotic organisms has evolved through time to form an integral part of ecosystems across our planet, shaping in ways both positive and negative the world in which we live.