29.4 THE STEM: TRANSPORT OF CARBOHYDRATES THROUGH PHLOEM
Much of the body of a vascular plant is devoted to the uptake and transport of the water required by leaves rather than to photosynthesis. In contrast, all cells of the algal ancestors of plants would have been capable of photosynthesis. Roots and stems contribute indirectly to photosynthesis but produce little or no carbohydrate themselves. Thus, although vascular plants are photosynthetic, a large part of their body must be supplied internally with food.
29.4.1 Carbohydrates are pushed through phloem by an osmotic pump.
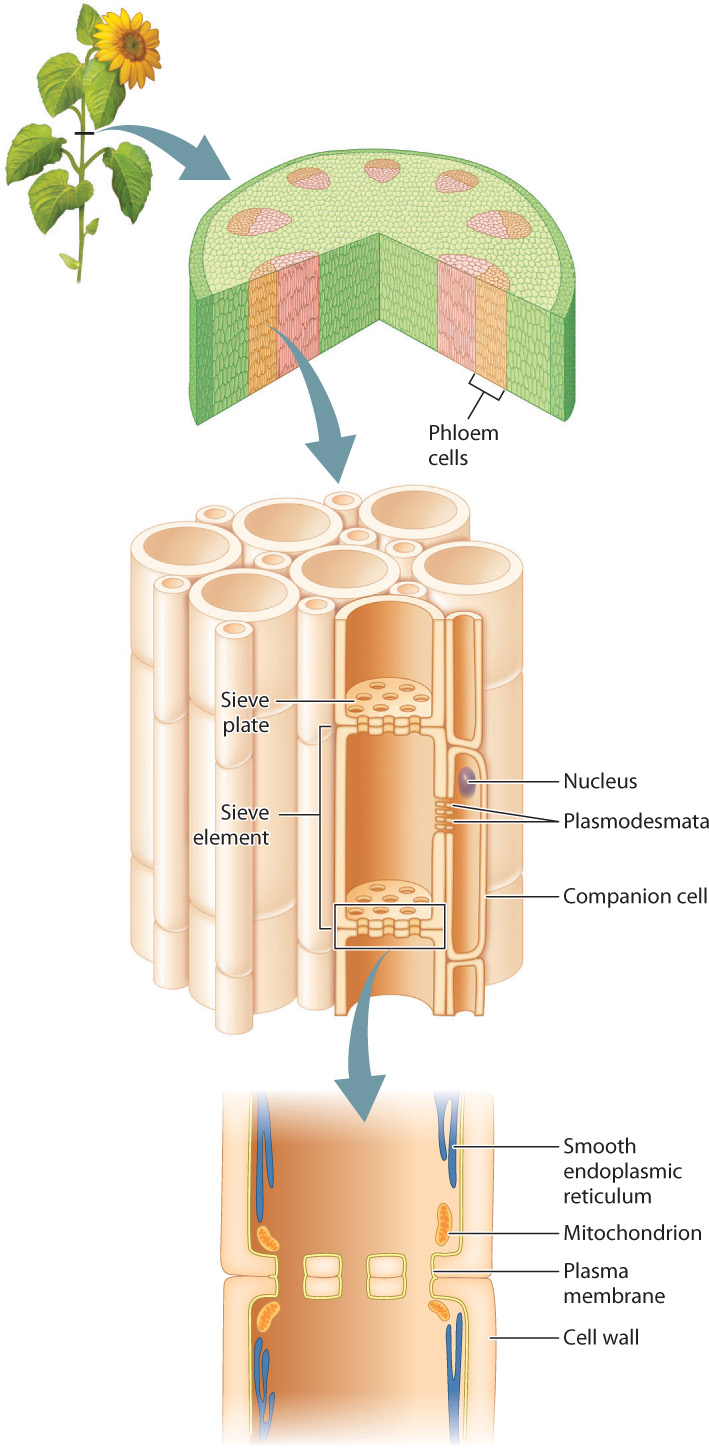
Phloem transport takes place through multicellular sieve tubes, which are composed of highly modified cells called sieve elements that are connected end to end (Fig. 29.15). During development, sieve elements lose much of their intracellular structure, including the nucleus and the vacuole. At maturity, sieve elements retain an intact plasma membrane that encloses a modified cytoplasm containing only smooth endoplasmic reticulum and a small number of organelles, including mitochondria. Cellular functions such as protein synthesis are carried out by an adjacent companion cell, to which the sieve element is connected by numerous plasmodesmata. Sieve elements are linked by sieve plates, which are modified end walls with large (1 to 1.5 μm diameter) pores. The plasma membrane of adjacent sieve elements is continuous through each of these pores, so each multicellular sieve tube can be considered a single cytoplasm-filled compartment. Phloem sap is the sugar-rich solution that flows through both the lumen of the sieve tubes and the sieve plate pores.
Phloem transports carbohydrates as sucrose (glucose plus fructose) or larger sugars, assembled from monosaccharides in the cytoplasm. Phloem also transports amino acids, inorganic forms of nitrogen, and ions including K+, which are present in much lower concentrations. Finally, phloem transports informational molecules such as hormones, protein signals, and even RNA. Thus, phloem forms a multicellular highway for the movement of raw materials and signaling molecules across the entire length of the plant.
Phloem transports its molecular cargo from source to sink (Fig. 29.16). In plants, sources are the regions that produce or store carbohydrates. For example, leaves are sources because they produce carbohydrates by photosynthesis, and potato tubers (the part we eat) are sources because they can supply stored carbohydrates to the rest of the plant body. Sinks are any portion of the plant that needs carbohydrates to fuel growth and respiration—examples are roots, young leaves, and developing fruits. Whereas the direction of xylem transport is always up toward the leaves, the direction of phloem transport can be either up or down, depending on where the source and sink are relative to each other.
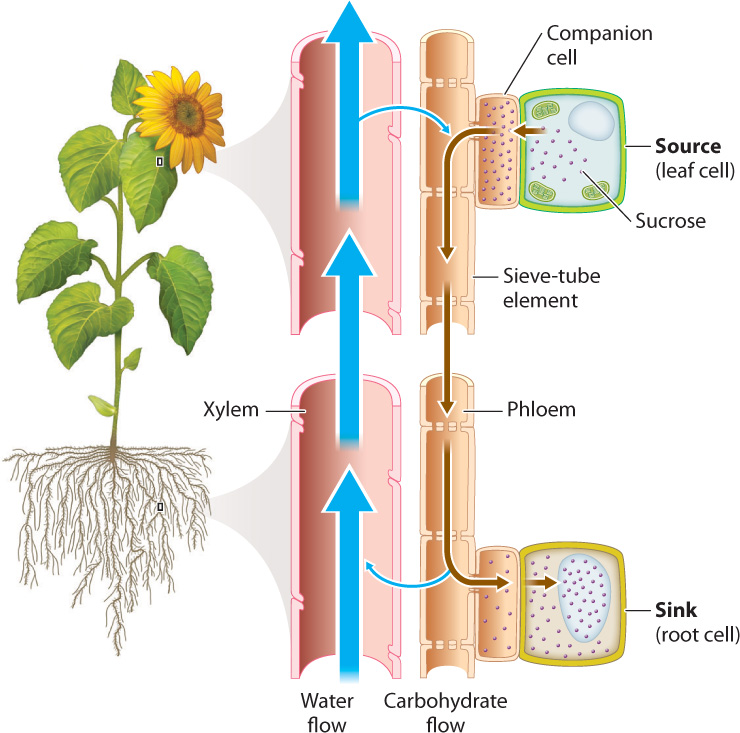
How does phloem transport sugars from source to sink? In some plants, active transporters powered by ATP move sucrose into phloem. The buildup of sugar concentration causes water to be drawn into the phloem by osmosis. Because the cell walls of the sieve tube resist being stretched outward to accommodate the influx of water, they press back (inward) against the cytoplasm. Their resistance to stretching increases the turgor pressure (Chapter 5) at the source end of a sieve tube. At sinks, sugars are transported out of the phloem into surrounding cells. This withdrawal of sugars causes water to leave the sieve tube, again by osmosis, reducing turgor pressure at the sink end. It is the difference in turgor pressure that drives the movement of phloem sap from source to sink.
The water that exits the phloem can be used locally to support the enlargement of sink cells or it can be carried back to the leaves in the xylem. Thus, some of the water in the phloem sap is recirculated in the xylem (Fig. 29.16). The volume of water that moves through the phloem, however, is tiny compared to the amount that must be transported through the xylem to replace water lost by transpiration. Therefore, the number and size of xylem conduits greatly exceeds the number and size of sieve tubes.
Phloem transport is sometimes referred to as “translocation,” a term that pairs well with “transpiration.” Yet in almost every way, phloem and xylem are a study in contrasts. In phloem, the plant generates the gradient that drives transport, whereas water moving through xylem is driven by the difference in hydration between soil and air. Furthermore, water is pulled upward through xylem conduits, while transport through phloem is more of a push.
These fundamental differences in function explain the cytological differences between xylem and phloem: Phloem conduits retain an intact plasma membrane and modified cytoplasm, whereas xylem conduits retain only their cell walls. What xylem and phloem have in common is that both are essential in enabling vascular plants to carry out photosynthesis on land. Moreover, like xylem, phloem is subject to risks that arise from the way flow through sieve tubes is generated.
Damaged sieve tubes are at risk of having their contents leak out, pushed out by high turgor pressures in the phloem. Damage is an ever-present danger because the sugar content of phloem makes it an attractive target for insects. Cell damage activates sealing mechanisms that repair breaks in sieve tubes. In some respects, these mechanisms are comparable to the formation of blood clots in humans, except that phloem can seal itself much more rapidly, typically in less than a second.
Quick Check 4
How is phloem able to transport carbohydrates from the shoot to the roots, as well as from the roots to the shoot (although not at the same time)?
29.4.2 Phloem feeds both the plant and the rhizosphere.
All the cells in a plant’s body contain mitochondria since all cells need a constant supply of ATP. Typically, about 50% of the carbohydrates produced by photosynthesis in one day are converted back to CO2 by respiration within 24 hours.
Carbohydrates that are not immediately consumed in respiration can be used as raw materials for growth, or they can be stored for later use. Carbohydrates stored within roots and stems, or in tubers (specialized storage organs such as potatoes), can support new growth in the spring or following a period of drought. Stored reserves can also be used to repair mechanical damage or replace leaves consumed by insects or grazing mammals. Like their green algal ancestors, vascular plants store carbohydrates primarily as starch, a large molecule that is not soluble and so does not affect the osmotic balance of cells.
What determines how carbohydrates become distributed within the plant? Where phloem sap ends up is determined by the sinks. Phloem transport to reproductive organs appears to have priority over movement to stems and leaves, and these have priority over movement to roots. In Chapter 31, we discuss the role that hormones play in controlling the growth and development of plants and how these hormones may influence the ability of different sinks to compete successfully for resources delivered by the phloem.
Phloem also supplies carbohydrates to organisms outside the plant. A fraction of the carbohydrates transported to the roots spills out into the rhizosphere, the soil layer that surrounds actively growing roots. This supply of carbohydrates stimulates the growth of soil microbes. As a result, the densities of microbial organisms near roots are much greater than in the rest of the soil. These soil bacteria are important decomposers of soil organic matter, which is rich in nutrients such as nitrogen and phosphorus. Thus, the release of carbohydrates by roots into the soil is thought to enhance the ability of roots to acquire nutrients from the soil.