29.5 THE ROOT: UPTAKE OF WATER AND NUTRIENTS FROM THE SOIL
As anyone who has dug a hole in a forest or a field can attest, plants make a lot of roots (Fig. 29.17). Laid end to end, the roots of a single corn plant would extend over 600 km. A major reason for such extensive investment belowground is that, with the important exception of CO2, everything that a vascular plant needs to build and sustain its body enters through its roots. This includes, of course, water, as well as nutrients such as nitrogen, potassium, and phosphorus.

Roots are an evolutionary innovation of the vascular plants; there were no roots when the ancestors of plants first moved onto land. The first roots would have encountered a land surface much less hospitable than today’s soils. To a large extent soils have a biological origin. The processes by which roots obtain nutrients from the soil accelerate rates of chemical weathering, stimulate the growth of microbes, and slow erosion, all of which contribute to the development of soils.
29.5.1 Nutrient uptake by roots is highly selective.
Xylem moves both water and nutrients (for example, nitrogen and phosphorus) from the root to the rest of the plant. Xylem, however, is never in direct contact with the soil. The living cells that surround the xylem allow roots to exert a high degree of selectivity over the movement of nutrients from the soil. By taking up only the nutrients they need, roots can concentrate certain nutrients; they can also prevent the entry of substances that could cause damage.
Like leaves and stems, roots have an outer cell layer of epidermis (Fig. 29.18). Epidermal cells in active areas of the root produce slender outgrowths known as root hairs. Root hairs require little material to build, but they greatly increase the surface area of the root available for absorbing nutrients. Inside the epidermis is the cortex, composed of parenchyma cells. Depending on species, the cortex can vary in thickness from a single cell to many cells. Xylem and phloem are located at the center of the root and are surrounded by the endodermis, a layer of cells that acts as a gatekeeper controlling the movement of nutrients into the xylem.
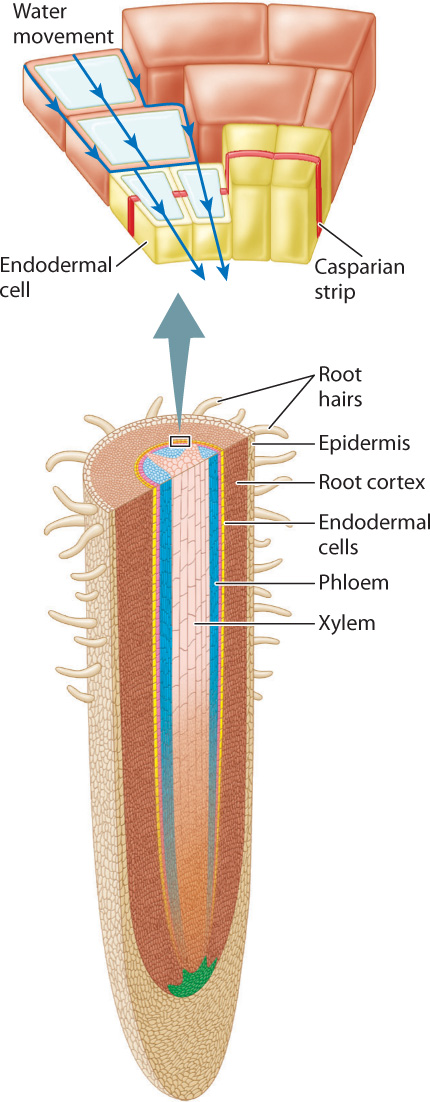
Water flowing across the epidermis and the root cortex can travel through the cells, crossing plasma membranes, and can also flow through the relatively porous walls that surround each cell. At the endodermis, flow through the cell wall is blocked by the Casparian strip, a thin band of hydrophobic material that encircles each cell. Water can gain access to the xylem only by passing through the cytoplasm of the endodermal cells.
The significance of the Casparian strip is that it allows the root to control which materials enter the xylem. How does it do that? In order for solutes to pass through the plasma membrane into the cytoplasm, they must be transported by ATP-powered proteins in the membranes of endodermal cells. These transporters move only certain compounds through the endodermis and actively exclude others. Thus, the solute composition of xylem sap—the fluid moving through the xylem—does not mirror the solute composition of water in the soil. By using the energy of ATP to control the movement of materials across the plasma membrane of endodermal cells, roots can concentrate nutrients that are needed in large amounts (for example, nitrogen) and exclude others that can damage plant cells (for example, sodium).
29.5.2 Nutrient uptake requires energy.
Roots expend a great deal of energy acquiring nutrients from the soil. We have already seen that energy is required to move nutrients across the cell membranes of the endodermis on their journey to the xylem. Energy is also required to make specific nutrients in the soil accessible to the root. And still more energy is required to support symbioses with soil microorganisms that take up nutrients and transfer them to plants. The mitochondria of root cells must, therefore, respire at high rates to provide energy needed for nutrient uptake. Their high respiration rates explain why plants are harmed by flooding: Waterlogged soils slow the diffusion of oxygen, limiting respiration.
The most abundant elements in the plant body—carbon, hydrogen, and oxygen—are obtained primarily from CO2 and H2O. All the other elements in the plant’s body come from the soil. To understand how roots acquire nutrients, let’s first look at how nutrients move within the soil.
Nutrients move through soil by diffusing through films of water that surround individual soil particles. In addition, when transpiration rates are high, the flow of water through the soil helps to convey nutrients toward the root. The uptake of nutrients by roots decreases their concentration at the root surface, creating a region of lower concentration toward which nutrients diffuse from the soil. However, because diffusion transports compounds efficiently only over short distances, each root is able to acquire nutrients only from a small volume of soil. This limitation explains why plants produce so many roots and so many root hairs: Both structures increase the volume of soil from which a root can obtain nutrients. It also explains why roots elongate more or less continuously, exploiting new regions of soil to obtain nutrients required for growth.
Nutrients vary dramatically in the ease and speed with which they move through the soil. For example, nitrate (NO3–) and sulfate (SO4–) are extremely mobile. In contrast, interactions with clay minerals keep zinc and inorganic phosphate (Pi) attached to mineral surfaces. One way roots gain access to highly immobile nutrients is by releasing protons or compounds that make the environment more acidic. Nutrients stuck to soil particles dissolve as water becomes acidic and diffuse to the root.
Soil microorganisms—both fungi and bacteria—are more adept than plants at obtaining nutrients. Fungi (Chapter 34) are particularly good at mobilizing phosphorus, and some bacteria can convert gaseous nitrogen (N2) into a chemical form that can be incorporated into proteins and nucleic acids (Chapter 26). What plants have at their disposal is a renewable source of carbohydrates. Using these carbohydrates, plants feed symbiotic fungi and bacteria, in effect exchanging carbon for phosphorus or nitrogen.
29.5.3 Mycorrhizae enhance the uptake of phosphorus.
Plants are able to access soil nutrients that their roots can’t reach by forming partnerships with fungi. Mycorrhizae are symbioses between roots and fungi that enhance nutrient uptake. Mycorrhizae are particularly important in enhancing the uptake of phosphorus, one of the most immobile nutrients in soil. The fungi absorb phosphate ions and transport some of them to root tissues; the fungi receive carbohydrates produced in leaves and transported to roots by the phloem.
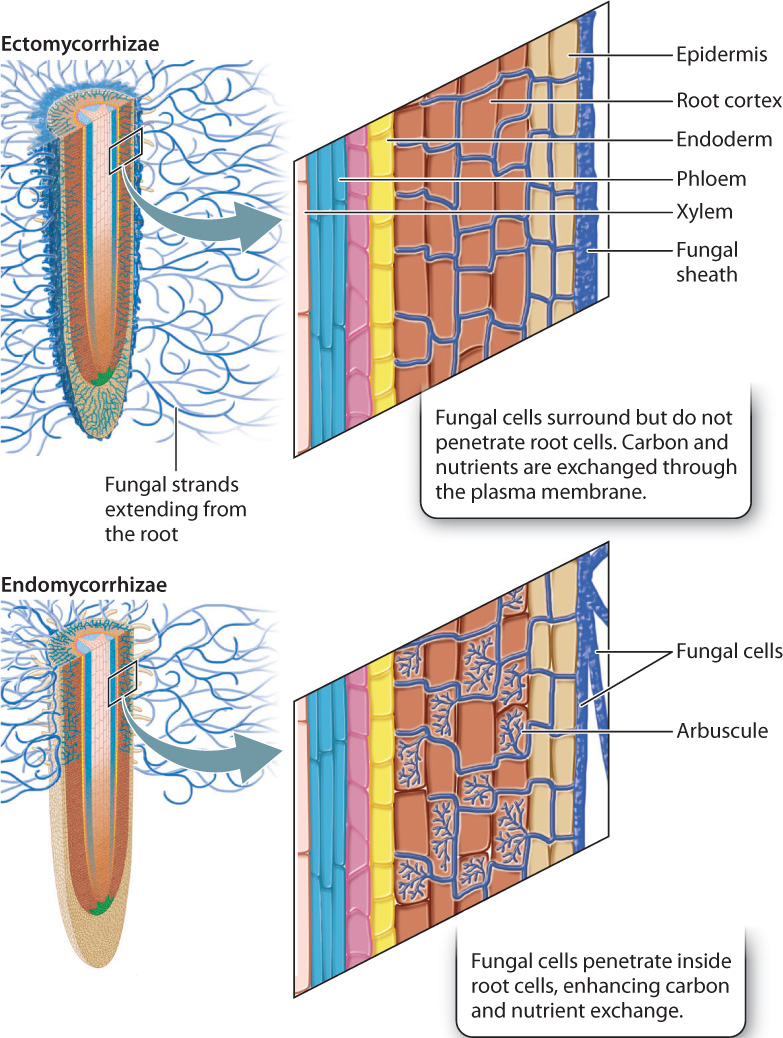
Why are fungi so effective at obtaining phosphorus from the soil? Roots are thin and root hairs thinner, but the cells produced by fungi are thinner still. Elongate fungal cells can extend more than 10 cm from the root surface, greatly increasing the absorptive surface area in contact with the soil by acting more or less like “super root hairs.” Mycorrhizae aid in nutrient uptake because solutes can be transported through the fungal cytoplasm much faster than they can diffuse through the soil. Furthermore, because fungal cells are so thin, they can penetrate small spaces between soil particles and thus access nutrients that would otherwise be unavailable. Finally, mycorrhizal fungi secrete enzymes that release phosphorus from soil organic matter and enhance the mobility of inorganic phosphorus in the soil.
Mycorrhizae are of two main types (Fig. 29.19). Ectomycorrhizae produce a thick sheath of fungal cells that surrounds the root tip. Some of these cells penetrate the walls of root cells, producing a dense network that surrounds individual root cells. The root and fungal cells are then able to exchange carbon and nutrients through their plasma membranes. Endomycorrhizae do not form structures that are visible on the outside of the root. Instead, they form highly branched structures, called arbuscules, that protrude into the interior of root cells. Despite this invasion, the plasma membranes of both cell types remain intact. The structure of the arbuscules increases the contact area between the two cell types, enhancing carbon and nutrient exchange.
Mycorrhizal associations are costly to the plant, consuming between 4% and 20% of its total carbohydrate production. When phosphorus is readily available from the soil, the percentage of mycorrhizal roots decreases, indicating some level of control by the plant. Nevertheless, mutualistic associations between roots and fungi occur in more than 85% of plant species. Mycorrhizal roots are thus the norm rather than an exception. Fossils from more than 400 million years ago suggest that interactions between plants and fungi may be ancient and perhaps helped plants gain a foothold on land.
29.5.4 Symbiotic nitrogen-fixing bacteria supply nitrogen to both plants and ecosystems.
Nitrogen is the nutrient that plants must acquire in greatest abundance from the soil, and in many ecosystems nitrogen is the nutrient that most limits growth. Nitrogen itself is not rare; 79% of Earth’s atmosphere is nitrogen gas (N2). However, as discussed in Chapter 26, plants and other eukaryotic organisms cannot use N2 directly. Only certain bacteria and archaeons have the metabolic ability to convert N2 into a biologically useful compound, a process known as nitrogen fixation. Nitrogen-fixing bacteria transform N2 into forms such as ammonia (NH3) that can be used to build proteins, nucleic acids, and other compounds. Some of these usable forms of nitrogen are released into the soil, making them available for use by plants and other organisms.
Although many nitrogen-fixing bacteria live freely in the soil, some live in plant roots. When the level of available soil nitrogen is low, some plants form symbiotic relationships with nitrogenfixing bacteria (Fig. 29.20). These interactions are highly species specific. In response to chemical signals transmitted between the root and bacteria, bacteria multiply within the rhizosphere. At the same time, root cells begin dividing, leading to the formation of a root nodule. Nitrogen-fixing bacteria enter the nodule either through root hairs or through breaks in the epidermis. Even though the bacteria appear to have taken up residence within the cytoplasm of nodule cells, in fact each one is contained within a membrane-bound vesicle produced by the host plant cell.
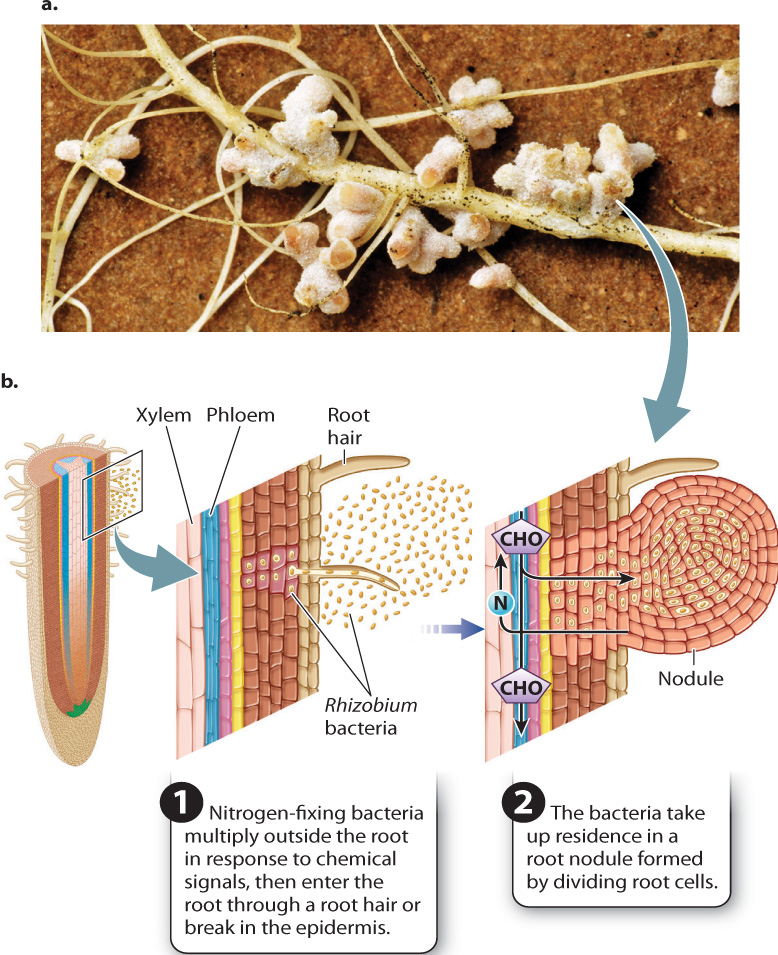
The plant supplies its nitrogen-fixing partners with carbohydrates transported by the phloem and carries away the products of nitrogen fixation in the xylem. As in mycorrhizal symbioses, the carbon cost to the plant is high: It is estimated that as much as 25% of these plants’ total carbohydrate production goes to nitrogen-fixing bacteria.
Relatively few plant species have evolved symbiotic associations with nitrogen-fixing bacteria, but those that do—especially members of the legume (bean) family—provide an important way in which biologically available nitrogen enters into ecosystems. In addition, nitrogen fixation in legume roots helps to sustain fertility in many agricultural systems.
Case 6 Agriculture: Feeding a Growing Population
29.5.5 How has nitrogen availability influenced agricultural productivity?
Harvesting crops removes nutrients—including nitrogen—from an ecosystem. These nutrients must be returned if the land is to to be agriculturally productive. In the early days of agriculture, the practice of shifting cultivation solved the problem by leaving fields unplanted for long periods between crops. During these periods, nitrogen-fixing bacteria replenished soil nitrogen, either in symbiotic association with roots or as free-living bacteria in the soil.
As populations grew, fields could not be left unplanted for long periods. Nutrients removed during harvest had to be replaced or the fertility of fields would decline. Organic fertilizers such as crop residues, manure, and even human feces provide a way to recyle other nutrients removed during harvest, but unless they are brought in from off the farm, they add little or no new nitrogen to the system.
Crop rotation has been widely practiced throughout the history of agriculture to resupply the soil with nitrogen. Every few years, the field is planted with legumes harboring nitrogen-fixing bacteria. Typically, the legume rotation is not harvested, but instead is treated as “green manure” that is allowed to decay and replenish soil nitrogen. During the eighteenth and nineteenth centuries, legume rotation contributed to the higher crop yields needed to support increasing populations. Therefore, along with the mechanization of agriculture, nitrogen-fixing symbioses contributed to the social conditions that fueled the Industrial Revolution.
By the second half of the nineteenth century, more abundant and more concentrated supplies of nitrogen fertilizers were needed to promote crop growth to feed growing populations. Mining of guano and sodium nitrate mineral deposits, both found in arid regions such as the northern deserts of Chile, helped supply the demand for the nitrogen needed to increase crop production. With time, however, the reserves of these natural nitrogen sources declined and a mechanism was needed to obtain nitrogen fertilizer from the vast storehouse of nitrogen in the atmosphere.
An industrial method of fixing nitrogen was developed by German chemist Fritz Haber in the years 1908–1911 and was subsequently adapted for industrial production by Karl Bosch. Their method allows humans to achieve what, until then, had been an entirely prokaryotic activity. Like nitrogen-fixing bacteria, the Haber–Bosch process requires large inputs of energy, but unlike bacteria, it occurs at high temperatures and pressures. The prospect of an essentially unlimited supply of nitrogen in a form that plants could make use of has proved irresistible. Industrial fertilizers produced by the Haber–Bosch process now account for more than 99% of fertilizer use. Industrial fixation of nitrogen today is approximately equal to the entire rate of natural fixation of nitrogen, which is largely due to bacteria, although a small amount of nitrogen is fixed by lightning.
As we discuss in Chapter 48, human use of fertilizer is altering the nitrogen cycle on a massive scale, posing a threat to biodiversity and the stability of natural ecosystems. Yet abundant nitrogen fertilizers are a cornerstone of the Green Revolution. The high-yielding varieties of wheat and rice introduced in the mid-twentieth century achieve their high levels of growth and grain production only when abundantly supplied with nitrogen. The fact that plants such as legumes (including beans, soybeans, alfalfa, and clover) form symbiotic relationships with nitrogen-fixing bacteria suggests a possible way around this problem. If it were possible to engineer nitrogen fixation into crops such as wheat and rice, either by inducing them to form symbiotic relationships with nitrogen-fixing bacteria or by transferring the genes that would enable them to fix nitrogen on their own, would that eliminate the need for nitrogen fertilizers? The answer is yes—but with a caveat. As we have seen, nitrogen-fixation requires a large amount of energy, diverting resources that could otherwise be used to support growth and reproduction.