30.3 TIMING OF REPRODUCTIVE EVENTS
As plants moved onto land, they encountered environments with distinct seasonality. In temperate regions, temperatures vary from summer to winter; in most tropical habitats, wet seasons alternate with dry ones. Thus, in addition to overcoming the challenges of completing their life cycle on land, plants evolved mechanisms to control the timing of reproduction. This is true for all plants, which must produce gametes at a time when they can find suitable mates and which must disperse their young in a season suitable for their growth.
However, timing is particularly important for angiosperms. Not only must individuals of the same species flower at the same time, but they must do so when they can attract enough pollinators for efficient pollination. Furthermore, angiosperms are unique among plants in including species with life-spans of only one to two years. For these short-lived species, which are most commonly found in temperate regions, there is little margin for error in the timing of flower production and seed germination. In this section, we explore how plants gain information about their environment that enables them to regulate when flowers are produced and seeds germinate.
30.3.1 Flowering time is affected by day length.
In the 1910s, Wightman Garner and Henry Allard, scientists at the U.S. Department of Agriculture, set out to understand why Maryland Mammoth, a variety of tobacco, does not produce flowers during the summer like other tobacco plants. Instead, these plants continue to grow throughout the summer, reaching heights several times that of a typical tobacco plant and producing many more leaves. Only in autumn does Maryland Mammoth finally produce flowers, too late in the year for seeds to mature.
One possible explanation for the observed delay in flowering time is that Maryland Mammoth must achieve a greater total size before it produces flowers. However, when Garner and Allard grew Maryland Mammoth plants in a heated greenhouse during the winter, the plants flowered at the same time and the same size as other tobacco plants. Therefore, size does not determine when Maryland Mammoth flowers. And because the heated greenhouse was set to the same temperatures as those found during the summer, temperature could also be eliminated as a factor. Garner and Allard were left with only one more environmental variable that was different between the summer and the winter: the length of the day. They proposed that plants can measure and respond to day length.
In a series of experiments in which day length was artificially shortened during summer and extended during winter, Garner and Allen showed that in Maryland Mammoth, as well as in many other plants, flowering is controlled by day length. They coined the term photoperiodism to describe the effect of the “photo period,” or day length, on flowering. Short-day plants, like Maryland Mammoth, flower only when the day length is less than a critical value. When the light period exceeds this threshold, a short-day plant continues to produce new leaves, but no flowers are formed. In contrast, long-day plants, which include radish, lettuce, and some varieties of wheat, flower only when the light period exceeds a certain length. Not all plants are photoperiodic. Flowering of day-neutral plants is independent of any change in day length.
Sensitivity to day length ensures that plants flower only when they are large enough to support the development of a large number of seeds and only when there is enough time for those seeds to mature before the onset of winter. A short-day plant that germinates in the spring will not flower until late summer, when the decreasing day length falls below the critical value for that species. This flowering strategy allows short-day plants to grow as large as possible and yet switch from producing leaves to producing flowers in time to complete seed development before winter. In contrast, many long-day plants germinate in the summer. Because they do not flower until a critical day length is exceeded, they do not flower in autumn, but instead during the following spring.
Understanding how photoperiod affects flowering has many commercial applications. For example, chrysanthemums are short-day plants and their natural flowering season is autumn. By artificially exposing plants to short day lengths, growers can induce chrysanthemums to flower at any time of the year. Photoperiodism can complicate the ability to grow crop species at latitudes different from the ones in which they evolved. For example, soybean is a short-day plant that normally germinates in the spring and flowers as the day length decreases in late summer. When planted in the tropics, where the day length is close to 12 hours throughout the year, soybean flowers when the plant is still quite small, reducing the number of seeds that can eventually be harvested. For this reason, crop species often become less sensitive to photoperiod as they are domesticated. For example, wild varieties of tobacco are photoperiodic, whereas cultivated tobacco, Nicotiana tabaccum, is day neutral.
30.3.2 Photoreceptors enable plants to measure day length.
The first requirement for measuring day length is that a plant be able to sense when it is day and when it is night. To accomplish this, many plants produce photoreceptors, molecules whose chemical properties are altered when they absorb light. We saw in Chapter 9 how a signal can be conveyed to a cell by a change in the shape of a receptor molecule within the cell or on its surface. Photoreceptors in plants work similarly. The absorption of light by a photoreceptor produces a signal that triggers changes in the cell’s metabolism or alters patterns of gene expression. Your eye relies on thousands of photoreceptors that work on the same principle as those in plants.
Although photoreceptors provide information about the presence of light, they do not, by themselves, explain how leaves are able to measure day length. An early clue to this ability came when scientists discovered that interrupting the dark period with a brief exposure to light was sufficient to alter the flowering response. In these experiments, both short-day plants and long-day plants were grown under a photoperiod that would normally cause the short-day plants to flower and the long-day plants to produce only leaves. A brief exposure to light during the dark period reversed this outcome. The short-day plants no longer produced flowers, but the long-day plants did. The reverse experiment, in which plants were placed under light-tight cover for a short period during the daytime, did not have any effect.
If no further experiments had been done, the scientists would have concluded— incorrectly—that plants were measuring the length of the dark period. More extensive experiments demonstrated that a plant’s sensitivity to a “night interruption” varies according to a circadian—or 24-hour—rhythm. We now know that photoperiodism results from an interaction between photoreceptors that are activated by light and the product of a gene whose expression is controlled by the plant’s circadian clock (Fig. 30.21). Circadian clocks are biochemical mechanisms found in all organisms that oscillate with a 24-hour period and are coordinated with the day-night cycle.
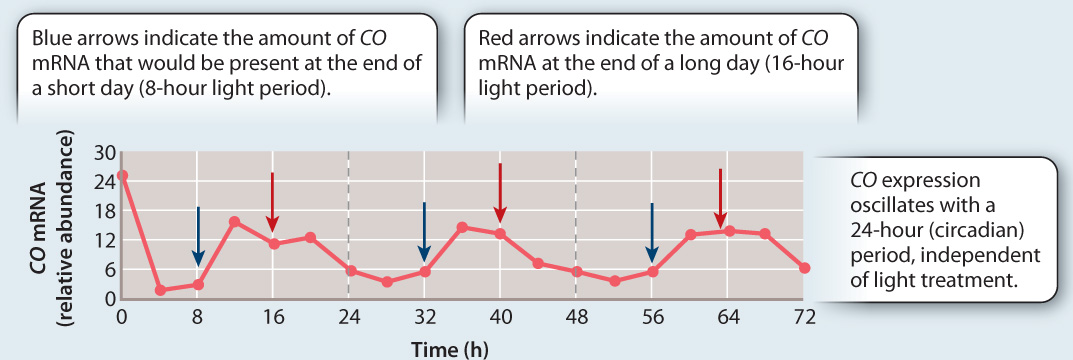
FIG. 30.21How do plants measure day length?
BACKGROUND Arabidopsis thaliana, a model organism for plant biology, is a long-day plant, flowering only when the light period exceeds about 16 hours. Genetic studies have shown that the CONSTANS (CO) gene is essential in allowing Arabidopsis to flower in response to day length. To determine the mechanism underlying photoperiodism, researchers set out to determine how this gene is affected by both the plant’s circadian clock and the presence or absence of light.
HYPOTHESIS The ability to respond to day length in Arabidopsis results from a circadian (daily) increase and decrease in CO expression, as well as interactions with light.
EXPERIMENT 1 The researchers determined how CO mRNA varies as a function of time of day and presence or absence of light.
RESULT 1 CO mRNA levels follow a circadian up-and-down pattern. This pattern occurred independent of whether the plant was exposed to long or short days or even kept continually in light. The timing of these oscillations is such that high levels of CO mRNA coincide with the light period only when day-lengths are long.
EXPERIMENT 2 Protein levels were measured following exposure to two days of light or dark. Protein extracts were first separated by gel electrophoresis and then probed with antibodies. Darker bands correspond to greater staining by protein-specific antibodies and thus provide visual information on the relative amount of the CO protein.
RESULT 2 The much darker band after exposure to two days of light indicates that CO protein accumulates only in the light. In the dark, the CO protein does not accumulate.

CONCLUSION Flowering in Arabidopsis is triggered by long days because there is overlap between high rates of CO expression and stabilization of the CO protein by light.
FOLLOW-UP WORK Studies with Arabidopsis mutants indicate a role for specific photoreceptors in stabilizing CO protein. Studies on other species indicate that photoperiod detection involves both the circadian clock and photoreceptors, but the way the CO protein is regulated by light varies.
SOURCES Suárez-López, P., K. Wheatley, F. Robson, H. Onouchi, F. Valverde, and G. Coupland. 2001. “CONSTANS Mediates Between the Circadian Clock and the Control of Flowering in Arabidopsis.” Nature 410:1116–1120; Valverde, F., A. Mouradov, W. Soppel, D. Ravenscroft, A. Samach, and G. Coupland. 2004. “Photoreceptor Regulation of CONSTANS Protein in Photoperiodic Flowering.” Science 303:1003–1006.
Other experiments demonstrated that day length is sensed by the leaves and not by the buds where the flowers will form. When conditions are right for flowering, a signal is transported from the leaves to the growing points of a plant. A flowering signal (named “florigen”) had been hypothesized to exist for more than 70 years, but its nature had not been known, although widely sought. Only with the development of techniques for genetic analysis were scientists able to investigate the genetic basis of photoperiodism and determine the identity of this important signal.
30.3.3 Vernalization prevents plants from flowering until winter has passed.
In a number of temperate-zone species, flowering can be induced only if the plant has experienced a prolonged period of cold temperatures, a process known as vernalization. Vernalization prevents plants from flowering prematurely in late summer or autumn. Plants are forced to wait until the following spring to flower, when the chances for successful pollination and seed development are greater.
An important feature of vernalization is that even parts of the plant that form after the cold stimulus is long past seem to “remember” that they received the required cold treatment. This “memory” of winter is important because many plants that overwinter grow rapidly in the spring. What is the basis of this “memory” of winter and how is it transmitted to new growth?
Plant species differ in the ways that vernalization affects the regulatory pathways that lead to flowering. These differences likely reflect the fact that the angiosperms evolved in tropical latitudes. As different groups migrated into temperate regions they independently evolved mechanisms for controlling flowering in highly seasonal environments.
One common mechanism acts through chromatin remodeling. The DNA of all eukaryotic organisms is bound with proteins to form chromatin. We saw in Chapter 19 that modifications to chromatin, such as DNA methylation, can change gene expression. By this means, a record of developmental events can be inscribed into a cell’s DNA. In Arabidopsis, for example, vernalization results in chromatin remodeling that turns off a gene whose protein product represses flowering. Chromatin remodeling is stable through mitotic divisions, explaining why newly formed parts of a plant “remember” winter. However, the slate is wiped clean during meiosis, and the requirement for vernalization is reinstated with each generation.
30.3.4 Dormant seeds can delay germination if they detect the presence of plants overhead.
Seed germination is a one-way street. Once a seed begins to germinate, it quickly reaches a point of no return and it either becomes established as a photosynthetically competent seedling or it dies. Dormant seeds may delay germination even though environmental conditions, notably temperature and moisture, are favorable. Dormancy increases the probability that some offspring will survive to maturity by extending the time over which the seeds germinate. From a farmer’s perspective, however, dormancy is a problem because it means that some fraction of the seeds put into the ground will not germinate in that year. Therefore, agriculturalists have paid close attention to what environmental factors induce germination.
In some species, dormancy is enforced by a hard, tough seed coat. In such species, the seed coat must be weakened mechanically before the seed will germinate, either by passing through an animal’s digestive system or by physical abrasion. In other plants, dormancy is controlled by the embryo. Two environmental variables are particularly effective at inducing such seeds to germinate. One is exposure to a prolonged period of cold temperatures, which signals that winter has past and thus prevents seeds from germinating on warm days in autumn. The second is that some seeds, particularly very small ones, will germinate only if they have been exposed to light.
When scientists first began to study the effects of light on seed germination they found that red light is particularly good at stimulating germination, while far-red light inhibits germination. A simple “flip-flop” experiment by H. A. Borthwick and colleagues showed that the inhibitory effect of far-red light could be overcome by a subsequent exposure to red light (Fig. 30.22). Ultimately it was shown that only a single photoreversible pigment was needed to detect the exposure to both far-red and red light.
FIG. 30.22How do seeds detect the presence of plants growing overhead?
BACKGROUND To study the effect of light on seed germination, scientists exposed lettuce seeds that had been kept continually in the dark to different wavelengths of light and then counted what fraction of the seeds germinated. Red light had the greatest ability to stimulate germination, but surprisingly far-red light inhibited germination such that fewer seeds germinated than in the control seeds, which were kept in darkness. In the rush to conduct more experiments, petri dishes with the light-treated seeds piled up by the sink until someone noticed that the seeds that had been experimentally treated with far-red light were now germinating.
HYPOTHESIS This observation suggested that the inhibitory effect of far-red light can be overcome by a subsequent exposure to red light.
EXPERIMENT The scientists exposed lettuce seeds to red and far-red light in an alternating pattern, ending with either red light or far-red light. They then placed the seeds in the dark for two days and afterward counted the number of seeds that had germinated.
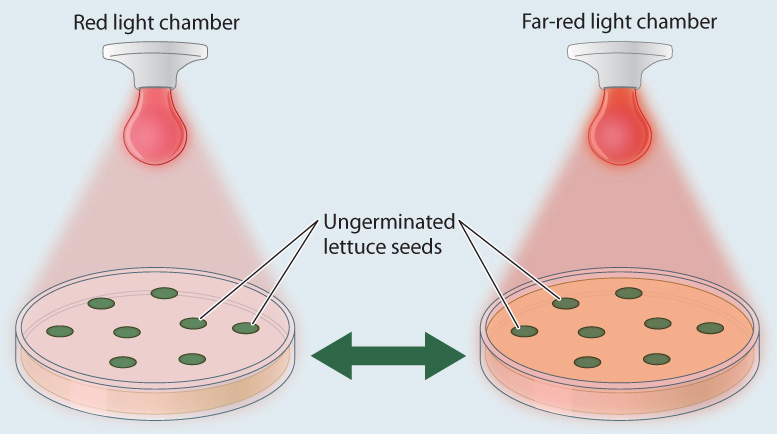
RESULTS When the lettuce seeds were exposed to red light last, nearly 100% of the seeds germinated. By contrast, when the last exposure of the seeds was to far-red light, the percentage of seeds that germinated was dramatically reduced.
| Lettuce Seeds Germinating After Exposure to Red and Far-red Light in Sequence | |||
|---|---|---|---|
| SEQUENCE OF LIGHT EXPOSURE | GERMINATION (%) | ||
| Dark (control) | 8.5 | ||
| Red | 98 | ||
| Red, Far-red | 54 | ||
| Red, Far-red, Red | 100 | ||
| Red, Far-red, Red, Far-red | 43 | ||
| Red, Far-red, Red, Far-red, Red | 99 | ||
| Red, Far-red, Red, Far-red, Red, Far-red | 54 | ||
| Red, Far-red, Red, Far-red, Red, Far-red, Red | 98 | ||
| TIME → | |||
CONCLUSION Seed germination in lettuce is triggered by exposure to red light and is inhibited by exposure to far-red light in a reversible fashion. As a result, plants are able to track changes in the relative amount of red and far-red light, which provides information on the presence or absence of plants overhead (see Fig. 30.23).
DETECTION OF SHADING BY PHYTOPCHROME Phytochrome detects the relative absence of red light under the canopy.
FOLLOW-UP WORK Additional studies showed that this result was due to a single pigment in a photoreceptor, now known as phytochrome, that is converted into an active form by red light and reversibly converted into an inactive form by far-red light.
SOURCE Borthwick, H. A., S. B. Hendricks, M. W. Parker, E. H. Toole, and V. K. Toole. 1952. “A Reversible Photoreaction Controlling Seed Germination.” Proceedings of the National Academy of Sciences 38:662–666.
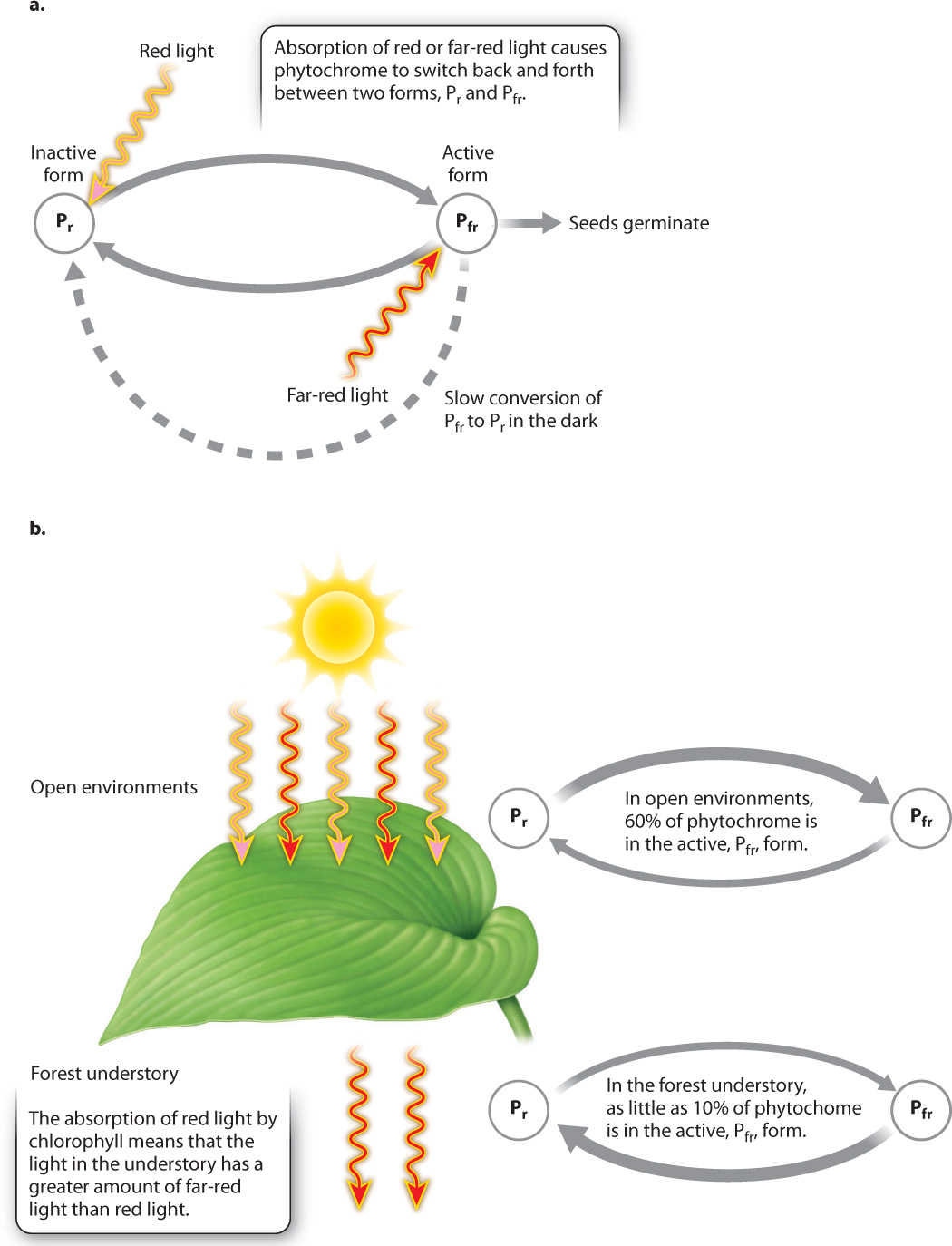
That pigment is present in phytochrome, a photoreceptor that switches back and forth between two stable forms depending on its exposure to light (Fig. 30.23a). Red light causes phytochrome to change into a form that absorbs primarily far-red light (Pfr); far-red light causes phytochrome to change back into the red-light-absorbing form (Pr). Because red light stimulates seed germination, we know that Pfr is the active form of phytochrome. In the dark, Pfr slowly converts back into the inactive, Pr, form. The light-independent conversion of Pfr to Pr allows a seed that had been exposed to red light, but failed to germinate, to have its phytochrome reset so that it can again respond to red light.
Phytochrome allows dormant seeds to respond to the presence of plants overhead. For a small seed that has few reserves, germinating in the shade of another plant could be fatal. How does phytochrome allow plants to detect the presence of other plants? As sunlight passes through leaves, the red wavelengths are absorbed by chlorophyll but the far-red wavelengths are not. The ratio of red to far-red light is thus much higher under an open sky than it is in the shade of another plant (Fig. 30.23b). As a result, the proportion of phytochrome that is in the active, Pfr, form is much greater in open environments than in the forest understory. Furthermore, the ratio of red to far-red light is independent of light intensity. Thus, the ratio is a reliable signal, even for seeds covered by a thin layer of soil, that leaves are overhead.
Phytochrome plays other important roles in plant development. As we will discuss in Chapter 31, phytochrome allows plants to alter their growth in the presence of neighboring plants to compete more eff ectively for sunlight. Phytochrome also maintains the accuracy of the plant’s circadian clock by resetting it each day at dawn. Finally, phytochrome is one of the photoreceptors that plants use to determine changes in day length.
Case 6 Agriculture: Feeding a Growing Population
30.3.5 What is the basis for the spectacular increases in the yield of cereal grains during the Green Revolution?
Mechanisms to control the timing of flower production and seed germination have evolved to optimize reproductive success under natural conditions. Agricultural conditions, however, are quite different. When Norman Borlaug, an American agronomist, led a project in the mid-twentieth century to make Mexico self-sufficient in bread wheat, his group encountered problems of flowering time and seed germination. Borlaug’s aim was to breed wheat for enhanced resistance to infection by a devastating class of fungal pathogens known as rusts. To do this, he and his group hand-pollinated plants that exhibited resistance to rusts with ones that grew well or had seeds that produced good flour.
Under natural conditions, wheat usually self-pollinates. To introduce new sources of pollen required that the anthers of each flower be removed by forceps before the flowers opened. Each flower had to be covered to prevent unwanted pollen from coming into contact with the flower. Finally, when the stigma was receptive, it had to be dusted with pollen collected from a plant with the desired characteristics. Under the hot sun, this is backbreaking work.
To speed the work, Borlaug proposed growing two crops of wheat each year. During summer, he conducted controlled crosses in the central highlands of Mexico, and then as winter approached he took his seeds north to the Yaqui Valley. There temperatures were warm enough for him to carry out another round of crosses during the winter.
Wheat is a long-day plant that flowers in the long days of summer. By successively growing plants during the lengthening days of summer in the southern highlands and the shorter days of winter in the lowland north, Borlaug was unconsciously selecting for plants whose flowering was less sensitive to photoperiod. Although not the original goal of the project, the diminishing sensitivity to photoperiod had an important impact. The new varieties were day neutral, flowering when they achieved a certain size. As a result, Borlaug’s varieties could be planted across a wide geographic range.
Before Borlaug’s rust-resistant seeds would be ready to be grown throughout the world, however, one more problem had to be solved. Having succeeded in breeding wheat varieties that were disease resistant, Mexican farmers applied fertilizer in large amounts. Under these conditions, the new wheat varieties produced so many seeds that stalks became top heavy, falling over in the wind.
Borlaug knew that an answer to this problem was to cross his plants with wheat varieties that had shorter, sturdier stems. But given the selective advantage of height in natural populations, where was he to find such a plant? Japanese wheat breeders had identified and preserved a mutant dwarf plant that occurred spontaneously, and Borlaug obtained 80 seeds of this dwarf variety. However, in his haste to incorporate this new plant into his breeding program, he made a serious error. He forgot that the Japanese wheats, which were adapted to a more northerly climate, had to be vernalized before they would flower.
Because the Japanese wheats had not been exposed to a prolonged period of cold, they did not flower at the same time as the Mexican wheats. And when they did produce a few flowers, the blooms were of such low quality that no viable seeds were produced. As luck would have it, Borlaug had planted only 72 of the 80 seeds. Somehow 8 seeds had escaped his notice. Eventually, Borlaug found these at the bottom of the original envelope in which they had been sent. This time he took care to vernalize the seeds, and when the new plants were formed they flowered readily.
Much hard work and many thousands of hand pollinations lay ahead. But by 1963, more than 95% of Mexican wheat cultivation made use of Borlaug’s high-yielding semidwarf wheat varieties, resulting in wheat yields six times greater than in 1944, the year he began work in Mexico. In 1965, the seeds were exported in large numbers, first to India and Pakistan and soon to the rest of the world. Used in combination with greater investments in fertilizer and irrigation, these varieties prevented the famines that had been predicted by many as a result of rising human populations. For his work in alleviating world hunger, Borlaug was awarded the Nobel Peace Prize in 1970.