31.3 SECONDARY GROWTH
Shoot apical meristems enable plants to grow in length and to branch. But a plant unable to grow in diameter would not become very tall before buckling under its own weight. And a plant unable to add vascular tissue could not supply its ever-increasing number of leaves with the water needed for photosynthesis. Growth in diameter thus serves two functions: It strengthens the stem, and it increases the capacity of the vascular system. Both are essential for supporting and supplying large numbers of leaves.
Primary growth is the increase in length made possible by apical meristems. An increase in diameter is referred to as secondary growth because it results from a new type of meristem, one that forms only after elongation below the growing tip is complete. The fossil record shows that the ability to grow in diameter evolved millions of years after primary growth, and it did so independently in several lineages, testimony to the benefits of being able to compete for sunlight by growing tall. Today, secondary growth occurs almost exclusively in seed plants, although this was not always the case in Earth history.
31.3.1 Shoots produce two types of lateral meristem.
Lateral meristems are the source of the new cells that allow plants to grow in diameter. Lateral meristems are similar to shoot apical meristems in their ability to produce new cells that grow and differentiate. Lateral meristems, however, differ from their apical counterparts in several key ways. First, unlike shoot apical meristems, which are located at the shoot tip, lateral meristems form cylinders or, more properly, cones of cells that extend the entire length of the stem. Second, because lateral meristems form only after elongation is complete, the new cells they produce grow in diameter but not in length. Finally, as a stem becomes thicker, the number of meristem cells needed to encircle the stem also increases. Thus, lateral meristems become larger over time.
Plants produce two distinct lateral meristems that together result in secondary growth. One of these, the vascular cambium (plural, cambia), is the source of new xylem and phloem. Secondary xylem is familiar to you: It’s wood. Plants with secondary growth are often described as woody, whereas plants that lack secondary growth and so are produced entirely by apical meristems are described as herbaceous. In a large tree, most of the plant’s mass consists of wood, generated from the vascular cambium.
As the activities of the vascular cambium increase the plant’s diameter, epidermis formed during primary growth eventually ruptures. Thus, plants with secondary growth have a second lateral meristem, the cork cambium, which renews and maintains an outer layer that protects the stem against herbivores, mechanical damage, desiccation, and, in some species, fire. These tissues (along with secondary phloem) are commonly known as bark. The word “cambium” derives from the Latin word cambiare, meaning “to change.” As we will see, the development of lateral meristems fundamentally changes the character of stems.
31.3.2 The vascular cambium produces secondary xylem and phloem.
The vascular cambium forms at the interface between xylem and phloem. In stems, the vascular cambium derives from procambial cells within each vascular bundle and parenchyma cells in between the bundles. In forming vascular cambium, both types of cell take on a new identity, becoming meristem cells.
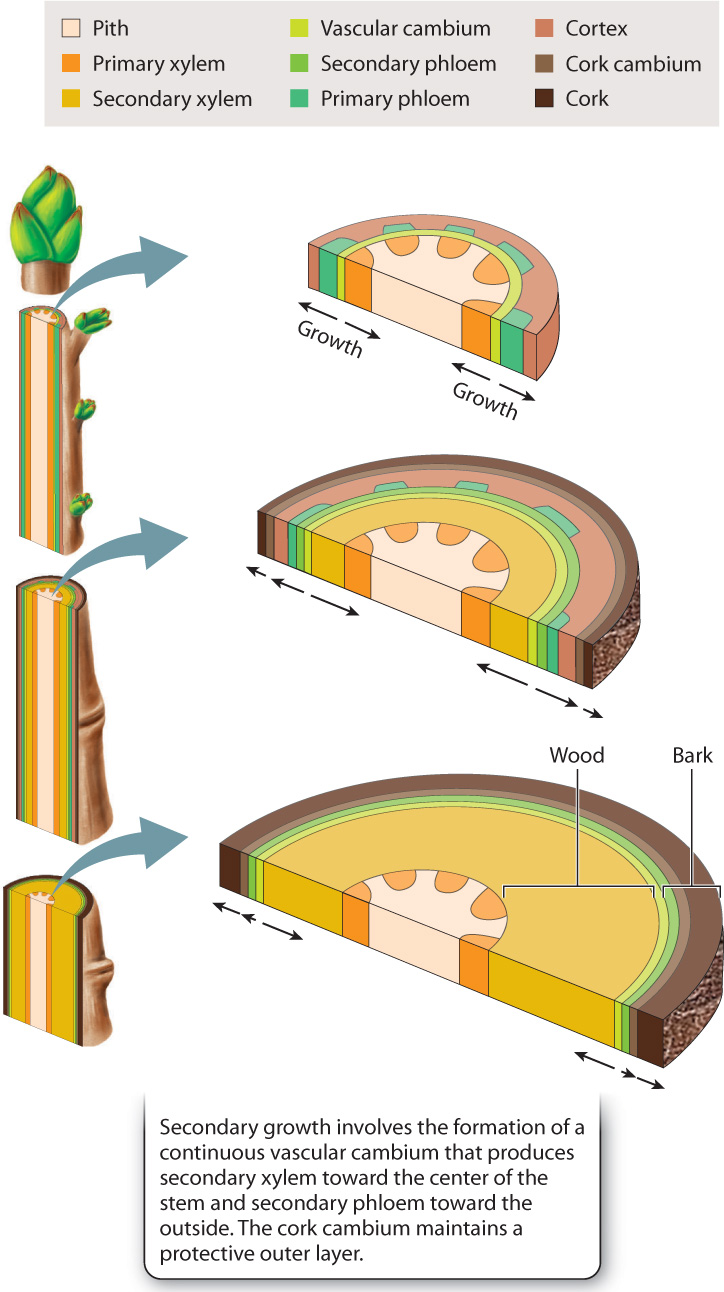
The vascular cambium produces new cells that differentiate on both its sides. Those to the inside of the vascular cambium become secondary xylem, and those on the outside develop as secondary phloem (Fig. 31.12). When you peel the bark off a stem, it separates from the wood at the vascular cambium. This is because the undifferentiated and actively dividing cells of the vascular cambium are weak and easily pulled apart. The vascular cambium forms a continuous layer, one to several cells thick, which surrounds the xylem and runs nearly the entire length of the plant.
The high rates of water loss by leaves associated with their uptake of CO2 for photosynthesis (Chapter 29) means that plants must produce more secondary xylem than secondary phloem. As the vascular cambium forms new secondary xylem cells along its inner face, the newly formed cells push the vascular cambium outward, much the way the elongation of newly formed cells pushes the shoot apical meristem upward. The vascular cambium accommodates this outward movement by adding new cells that increase its circumference. In contrast, the secondary phloem, once formed, is unable to produce any additional cells. As the production of secondary xylem pushes the vascular cambium outward, the phloem becomes stretched and, eventually, damaged. Consequently, new secondary phloem must be continually produced.
Over time, xylem conduits lose their ability to transport water. Thus, in long-lived trees, the functional xylem is located in a sapwood layer adjacent to the vascular cambium, while the heartwood, in the center of the stem, does not conduct water. Heartwood is often darker in color than sapwood because it contains large amounts of chemicals such as resins. For this reason, heartwood is typically more resistant to decay than sapwood.
In many plants in seasonal climates, decreases in the size of secondary xylem cells at the end of the growing season result in growth rings, which make it possible to determine a tree’s age. Growth rings also provide a window into the past because by measuring the width of each ring, it is possible to determine how much wood was laid down in a given year (Fig. 31.13). Wood production is higher when the environmental conditions that enable photosynthesis are more favorable. Thus, annual growth rings provide a record of past climatic conditions such as temperature and moisture, as well as events such as fire.

31.3.3 The cork cambium produces an outer protective layer.
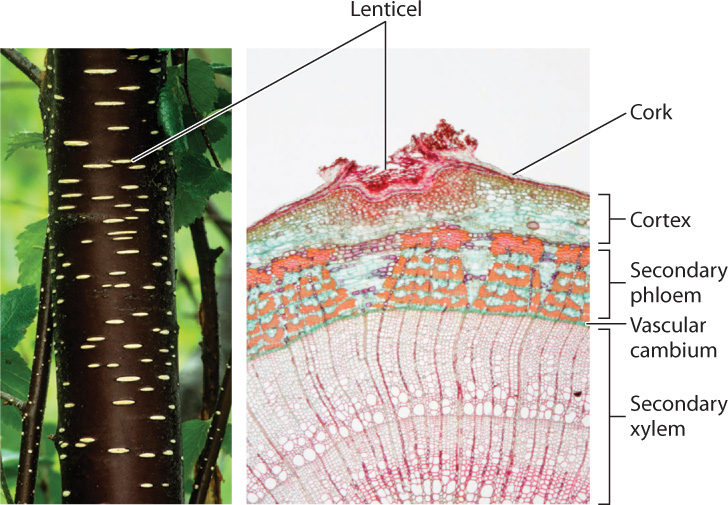
The role of the cork cambium is to maintain a protective layer around a stem that is actively increasing in diameter (see Fig. 31.12). The cork cambium forms initially from cortex cells that dedifferentiate—that is, they regress to their earlier state—to become meristem cells. However, with time, this layer of actively dividing cells becomes increasingly cut off from its source of carbohydrates—the phloem. When the cork cambium becomes sufficiently cut off from its carbohydrate supply, a new cork cambium forms within the secondary phloem. Thus, each cork cambium is relatively short lived, and in many cases, new cork cambia form in patches rather than as continuous layers. As a result, the bark of many trees has a patchy and fissured appearance (Fig. 31.14).

The cells produced by the cork cambium are called cork. As cork cells mature, they become coated with suberin, a waxy compound that protects against mechanical damage and the entry of pathogens, while also forming a barrier to water loss. However, this layer of waxy, nonliving cells impedes the inward diffusion of oxygen needed to meet the respiratory demands of living cells. The outer bark cells are less tightly packed in regions called lenticels, allowing oxygen to diffuse into the stem (Fig. 31.15).
Quick Check 2
Why do plants have two types of lateral meristem?
31.3.4 Wood has both mechanical and transport functions.
We have seen that as a tree grows, its secondary xylem fulfills the two roles of secondary growth. The first is to provide trees with the strength and stability to stand upright and support large crowns. The second is to transport water and nutrients from the roots to the leaves. In most gymnosperms, a single cell type fulfills both functions. Cells on the xylem side of the vascular cambium differentiate to form tracheids (Fig. 31.16a). As discussed in Chapter 29, tracheids are single-celled xylem conduits that have thick lignified cell walls and are nonliving and empty at maturity. The wood of a pine tree, for example, is relatively homogeneous in structure, consisting almost entirely of tracheids. This type of wood does not vary dramatically from species to species.

Angiosperms, by contrast, exhibit a great diversity in wood structure, as the cells mature into several distinct types that separate the functions of water conduction and mechanical support (Fig. 31.16b). Fibers are narrow cells with extremely thick walls and almost no lumen; their primary function is support. In contrast, vessel elements can be extremely wide cells; their primary function is water transport (Chapter 29). The largest vessel elements are close to 0.5 mm in diameter and thus visible to the naked eye. Vessel elements can be much wider than tracheids because the fibers provide support. And because vessel elements develop open connections that link them end to end with other vessel elements, creating multicellular xylem vessels, they allow angiosperms to produce wood that is more efficient at transporting water than the wood of gymnosperms.
The ability to form xylem vessels has a number of important consequences. One of these is that angiosperms can have higher rates of water flow through their stems than is possible with wood that contains only tracheids. As a result, angiosperms sustain higher rates of CO2 uptake and thus higher growth rates. In addition, because vessels can be much more efficient than tracheids at transporting water, angiosperms can devote a smaller fraction of their wood to water conduction, and more of their stem volume to other cell types. The wood of some angiosperms contains many fibers, in extreme cases resulting in wood that is so dense that it sinks. Such dense woods are extremely strong, allowing trees to create spreading crowns with outstretched branches. Such “hardwoods” are useful as flooring materials and furniture. The wood of other angiosperms contains a high percentage of living cells instead of fibers, and this wood has a very low density. Balsa trees, whose light wood is used in building model airplanes, are a good example.