31.4 ROOT GROWTH AND DEVELOPMENT
The roots that evolved in vascular plants enabled them to obtain water and nutrients from the soil and provided a mechanical anchor for plants capable of growing tall (Chapter 29). As in the case of leaves and wood, the fossil record shows that the first vascular plants lacked roots. These plants would have been limited to extremely wet environments in which they could absorb water through horizontal, and possibly submerged, stems. The evolution of roots, then, allowed vascular plants to expand into drier habitats and ensured the stability of taller stems.
The fossil record also indicates that roots evolved independently in the lycophytes and the lineage leading to the other vascular plants. Nevertheless, the roots of all vascular plants share many features. This suggests that the physical challenges of growing through a dense medium such as soil and the demands of obtaining water and nutrients from that medium place strong constraints on root structure. Here, we explore the growth and development of the plant body belowground, emphasizing how the elongation and branching of roots enables vascular plants to obtain the water and nutrients they need from the soil.
31.4.1 Roots grow by producing new cells at their tips.
In Chapter 29, we saw that plants produce large numbers of thin roots to obtain sufficient nutrients from the soil. And to obtain water during periods without rain, plants produce roots that penetrate deep into the soil. Growing roots into the soil presents challenges different from those associated with elongating stems into the air. Although the dangers of drying out are much reduced belowground, deeper soil layers can have densities approaching that of concrete. How are the thin and permeable roots needed for water and nutrient uptake able to penetrate and grow through the soil?
At one level, roots grow in the same way as shoots, with new cells produced at their tip. The root apical meristem serves as the source of new cells. After cell division, these cells undergo elongation and complete their differentiation only after elongation has ceased (Fig. 31.17). Thus, as we move back from the elongating root tip, we encounter successive regions of cell division, elongation, and maturation, just as we saw in stems.
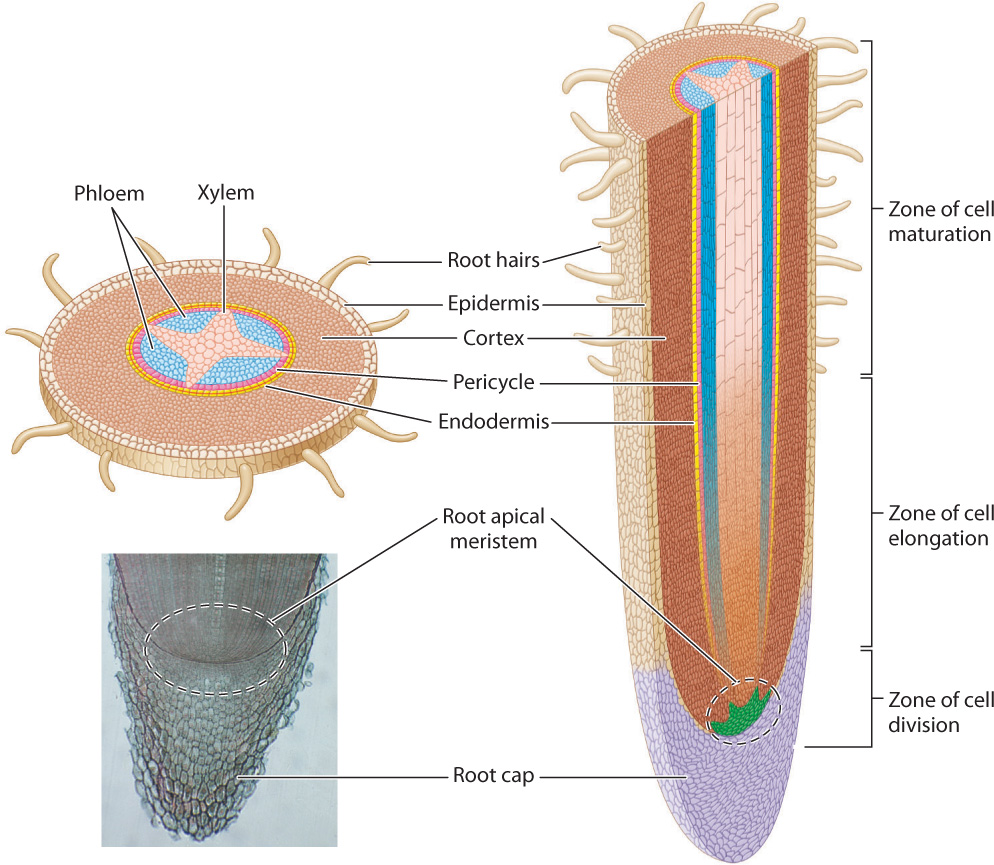
Roots, however, differ from stems in several important ways. One important difference is that roots are much thinner than stems. Plants with thin roots can more efficiently produce the large surface area needed for optimal uptake of water and nutrients from the soil. Thin roots can also more easily bend around soil particles and make use of channels created by invertebrate animals that burrow through the soil. A second major difference is that the root apical meristem is covered by a root cap that protects the meristem as it grows through the soil. As the root elongates through the soil, root cap cells are rubbed off or damaged. Thus, the root cap is continually supplied with new cells produced by the root apical meristem. A third difference between roots and stems, and one that can be seen without a microscope, is that the root apical meristem does not produce any lateral organs, whereas the shoot apical meristem makes leaves. Root hairs, the single-celled outgrowths of root epidermal cells that greatly increase the surface area for uptake (Chapter 29), are formed only after elongation has ceased.
Let’s now look at how roots develop the specialized cells and tissues that they need to function. A cross section taken through the zone of cell maturation shows a series of concentric tissue layers, starting with the epidermis on the outside, followed by the root cortex, and then the endodermis (Fig. 31.17). Recall that the endodermis plays a key role in the selective uptake of nutrients (Chapter 29). The Casparian strip, a waxy band surrounding each endodermal cell, forces all materials that will enter the xylem to cross the plasma membranes of these cells.
At the center of the root lies a single vascular bundle in which the xylem is arranged as a fluted cylinder, with phloem nestled in each groove. In cross section, the xylem is arranged in a star shape, the number of points of the star differing in different plant species.
As in stems, the hormone auxin plays a central role in specifying which cells differentiate into xylem and phloem. Auxin produced by developing leaves is transported downward through the mature stem and into the roots by the phloem. At the end of the differentiated phloem, auxin moves by polar transport from cell to cell until it reaches the root apical meristem and the root cap. As auxin moves through the elongation zone, it triggers the formation of procambial cells that subsequently differentiate into phloem and xylem.
In this way, the production of auxin by developing leaves and the polar transport of auxin from cell to cell ensures that vascular continuity is established between roots and leaves. Phloem extends closer to the root tip than do fully lignified xylem conduits. This reflects the fact that the apical meristem can obtain water and nutrients directly from the soil but depends entirely on the leaves for carbohydrates needed for growth.
So far, we have described only the primary growth of roots. However, roots can undergo secondary growth as well. This allows roots to increase in diameter and to add new xylem and phloem. As in stems, the vascular cambium forms at the interface between the xylem and the phloem. Indeed, the vascular cambium must be continuous from the shoot to the roots to ensure that the secondary xylem and phloem provide an unbroken connection between the roots and the leaves. In older roots with extensive secondary growth, a cork cambium is also formed. As in stems, the cork cambium renews the outer layers of cells that protect the living tissues from damage and predation.
31.4.2 The formation of new root apical meristems allows roots to branch.
Compared to stems, roots branch like crazy. Even a plant that has only a single stem aboveground will have many tens of thousands of elongating root tips. A major reason that roots branch so much is that the uptake of water and nutrients is concentrated in a zone near the root tip where root hairs are most abundant. Thus, branching is necessary to create enough root surface area to supply the water and nutrient needs of the leaves.
For a root to branch, a new root apical meristem must be formed. In roots, new meristems develop from the pericycle, a single layer of cells just to the inside of the endodermis (Fig. 31.18). The pericycle is immediately adjacent to the vascular bundle, so the new root is connected to the vascular system right from the beginning. But because the new root meristem develops internally, the new root must grow through the endodermis, cortex, and epidermis before reaching the soil.
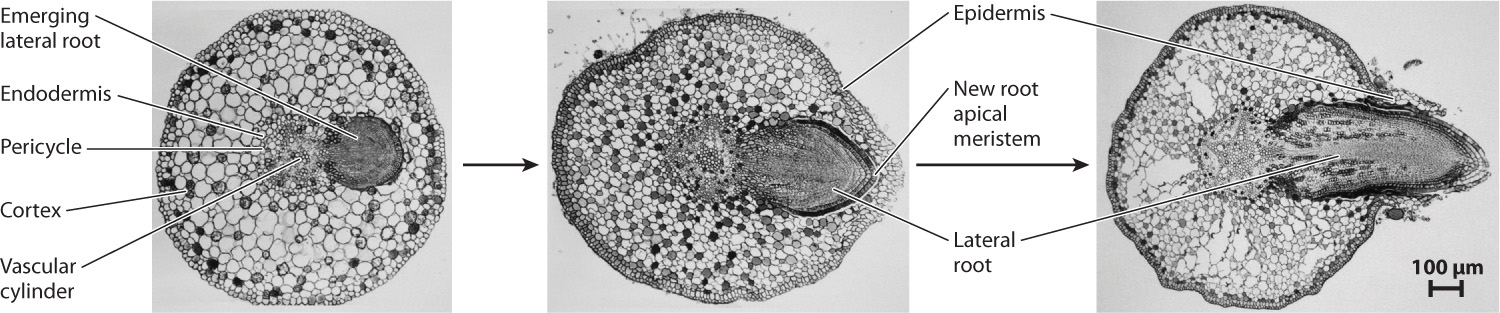
One consequence of initiating new root meristems from the pericycle is that new roots can form anywhere along the root. As a result, plants have tremendous flexibility in the development of their root systems and roots can proliferate locally in response to nutrient abundance. For example, nitrate (NO3–), the most common form of nitrogen in soil, triggers the formation of lateral roots by causing auxin to accumulate locally within the root tissues nearest to the nitrate-rich soil. Only a relatively small number of a plant’s roots, however, continue to elongate and branch indefinitely. Most new roots are active for only a short time, after which they die and are shed. This turnover of fine roots allows the root system to respond dynamically to changes in the availability of water and nutrients but represents a major expenditure of carbon and energy for the plant.
Up to this point, we have described how new roots form from existing roots. New roots can also be produced by stems. This is important in plants that lack secondary growth, many of which produce horizontal stems that grow on or beneath the soil surface, for example many lycophytes and ferns. In these plants, new roots form at each successive node, so the uptake capacity of the root system is directly coupled to the elongation of the stem.
When the stem of a plant is severed—and if the shoot tip is not allowed to dehydrate—new roots often form at the cut end. The new root meristems are stimulated by auxin, which is produced by the young leaves and accumulates, by polar transport, at the cut end. The ability of cut stems to form new roots allows plants to survive and in some cases proliferate following mechanical damage. It is also the means by which many commercially important plants are propagated.
Quick Check 3
How is root development similar to and how is it different from stem development?
31.4.3 The structures and functions of root systems are diverse.
In the preceding sections, we focused on the growth and branching of individual roots. How a plant’s roots are distributed in the soil has a tremendous impact on the ability of the root system to supply the shoot with water. Some plants produce very deep roots that provide them with access to a groundwater supply that is independent of the variability of rainfall. This can be advantageous, but it comes at the cost of producing and maintaining these non-photosynthetic structures. Roots can penetrate into mine shafts and caves more than 50 m below the soil surface.
At the other extreme are plants that produce shallow and spreading root systems. For example, barrel cacti produce roots that penetrate less than 10 cm into the soil but extend out several meters from the base of the plant. These roots cast a wide but shallow net to capture as much water as possible from intermittent rain showers.

Some plants produce distinctive roots whose principal role is other than the absorption of water and nutrients from the soil (Fig. 31.19). For example, climbing plants such as poison ivy produce roots along their stems that allow them to adhere to the sides of trees (Fig. 31.19a). The conspicuous prop roots produced by some tropical trees provide mechanical stability that allows these plants to grow tall despite their slender stems (Fig. 31.19b).
Many plants produce swollen roots in which resources such as starch can be stored (Fig. 31.19c). Belowground storage allows plants to produce new leaves and shoots quickly following either damage or a period in which drought or cold has caused the shoots to die. Cassava and sweet potato are examples of economically important species with storage roots.
Perhaps the most unusual rooting structures are the “breathing” roots produced by trees that grow in water. For example, black mangroves grow along the seashore in tropical regions and produce pencil-sized roots that extend vertically upward out of the sandy soil (Fig. 31.19d). Breathing roots do not actually breathe; instead, they contain internal air spaces that provide an easier pathway for oxygen to diffuse into the roots than the waterlogged soil. For this reason, breathing roots must extend above the surface of the water. Thus, the breathing roots of black mangrove provide an estimate of the water level at high tide.