32.2 DEFENSE AGAINST HERBIVORES
The first land plants were followed closely in time by the ancestors of spiders and scorpions. Fossils show that these earliest land animals soon began to feed on plant fluids and tissues. As plants diversified, so did herbivores, and the pressure on plants to defend themselves against organisms from caterpillars to cows only increased. Given that they are unable to run away, plants seem like they would be the ideal food. But plants have evolved a diversity of mechanisms that deter would-be consumers. Because these mechanisms use resources that could otherwise be directed toward growth and reproduction, plants have also evolved means of deploying their defenses in a cost-effective manner. Here, we explore the mechanical, chemical, developmental, and even ecological means by which plants protect themselves in a world teeming with hungry herbivores.
32.2.1 Plants use mechanical and chemical defenses to avoid being eaten.
Milkweeds (Asclepias species) are commonly found in open fields and roadsides across North America. They illustrate well how plants protect themselves from herbivorous animals. In fact, most generalist herbivores, which eat a diversity of plants, do not use milkweeds as a food source. Only specialists, such as caterpillars of the monarch butterfly (Danaus plexippus), can feed on these well-defended plants.
In part, milkweed’s defenses are mechanical. The leaves of many milkweed species are covered with dense hairs. An insect crawling on a leaf must make its way through as many as 3000 hairs per square centimeter, a journey likely more daunting to an insect than wading into a blackberry thicket is to a human. Monarch caterpillars may spend as much as an hour mowing off these hairs before they begin to feed. This long handling time represents a significant cost. To understand why monarch caterpillars make this investment, let’s look at milkweed’s other defenses.
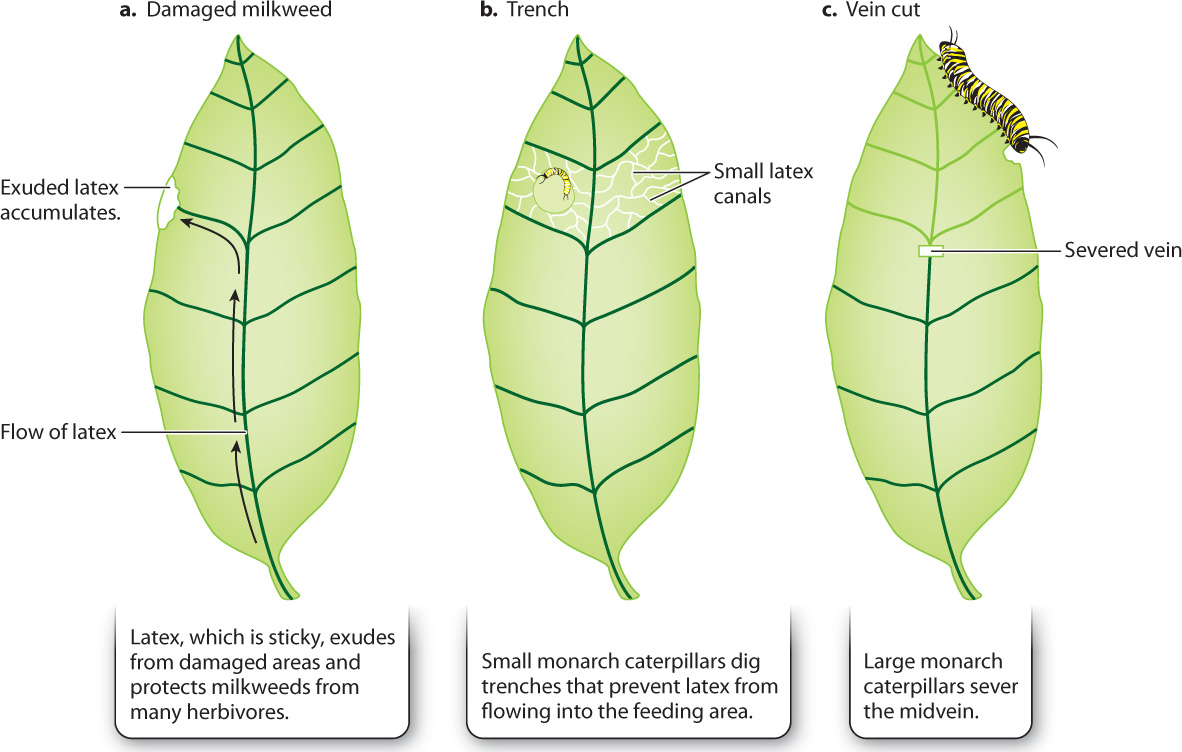
A network of extracellular canals filled with a white sticky liquid called latex runs through the milkweed leaf, both within the major veins and extending into the regions between veins (Fig. 32.10). Latex canals are under pressure in the intact leaf. Thus, when the leaf is damaged, the latex flows out, sticking to everything it comes into contact with and rapidly congealing on exposure to air. A feeding insect is in danger of having its tiny mouthparts glued together, with potentially fatal consequences. Small monarch caterpillars cut trenches into the leaf that prevent latex from flowing onto the region where they are feeding. Larger caterpillars sever the midvein, relieving the pressure within the latex canals downstream of the cut. The caterpillars are then able to feed on the part of the leaf that lies beyond the severed midvein without risk.
Milkweed latex is not only sticky but also toxic. In particular, milkweed latex contains high concentrations of cardenolides, steroid compounds that cause heart arrest in animals. (In small doses, these compounds are used to treat irregular heart rhythms in humans.) Monarch caterpillars do not metabolize the cardenolides that they consume, but instead sequester them within specific regions of their bodies. Storing these compounds provides the monarchs with a chemical defense against their own predators. Because of the presence of these cardenolides, monarch caterpillars and even the adult butterflies are highly toxic to most birds and other animals that would otherwise eat them.
Most plant species have mechanical or chemical defenses against herbivores, or even both. Hairs are common on leaves, and, as anyone who has ever rubbed against a stinging nettle knows, these hairs are sometimes armed with chemical irritants. Grasses and some other plants have a hard, mineral defense consisting of silica (SiO2) plates formed within epidermal cells. Silica functions as a mechanical defense because it wears down insect mouthparts, so the insects feed less efficiently and grow more slowly. On a larger scale, some tropical trees have prickles, spines, or thorns (Fig. 32.11). Even something as simple as how tough a leaf is can have a large impact on the probability of the leaf’s being eaten. In fact, leaves are most likely to be eaten while they are still growing and their tissues not fully hardened. Nonetheless, the most spectacular diversity in plant defenses lies in the realm of chemistry.

32.2.2 Diverse chemical compounds deter herbivores.
The cardenolides produced by milkweeds are only one example of the vast chemical arsenal available to plants for protection. Some plants produce alkaloids, nitrogen-bearing compounds that damage the nervous system of animals. Commonly bitter tasting, alkaloids include such well-known compounds as nicotine, caffeine, morphine, theobromine (found in chocolate), quinine (a treatment for malaria), strychnine, and atropine. Alkaloids are a costly defense because they are rich in nitrogen, an essential and often limiting element that plants need to build proteins for photosynthesis. However, they are specific in their action and therefore very small concentrations are effective. One of the first chemotherapy drugs (Table 32.1), vincristine, is an alkaloid extracted from the Madagascar periwinkle. It inhibits microtubule polymerization and thus prevents cell division (Chapter 11). Vincristine does not harm the plant’s own cells because it is stored within an extracellular system of latex-containing canals.
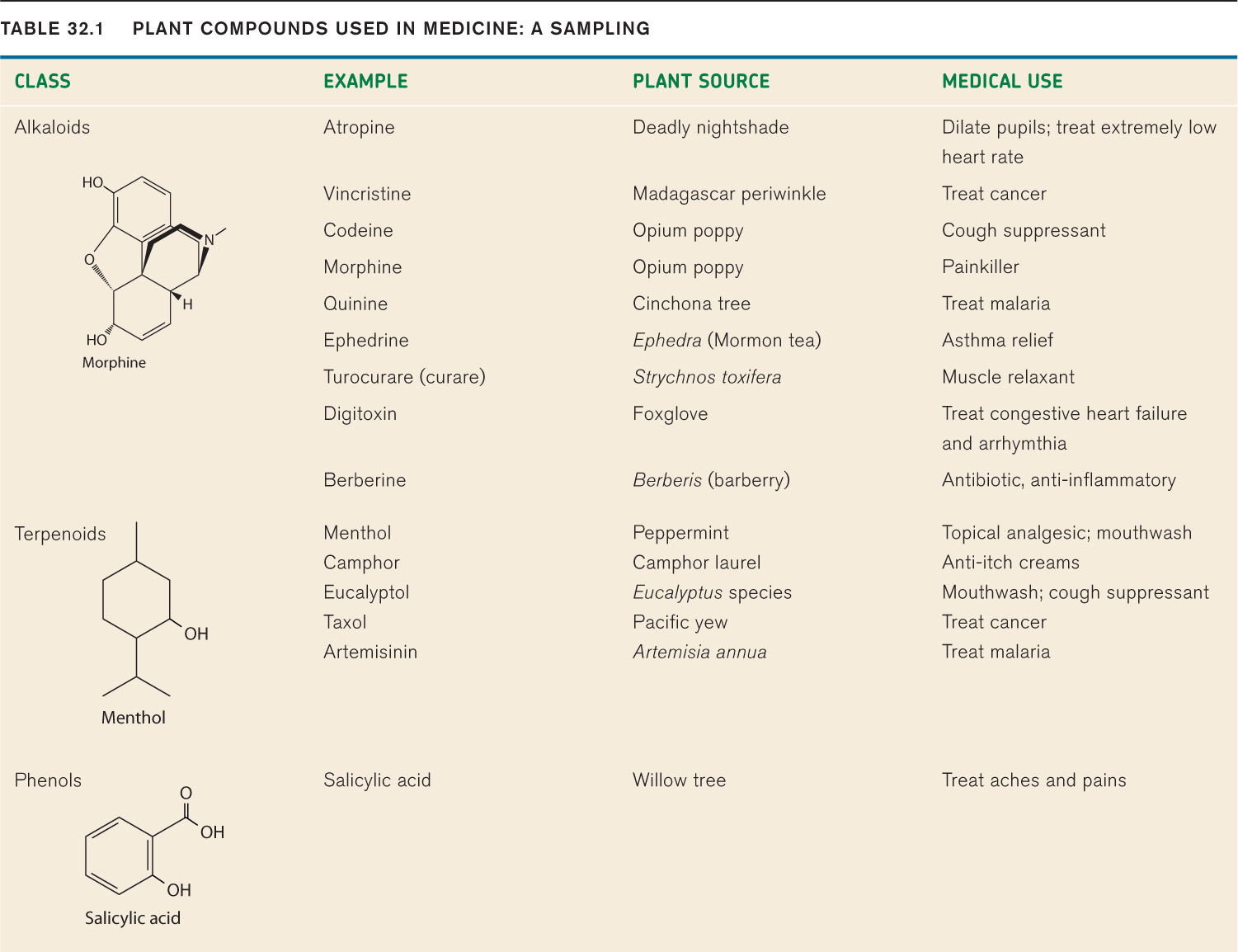
A second group of defensive compounds, the terpenes, do not contain nitrogen. As a result, plants can produce them for defense without having to make use of nitrogen that could otherwise be used in protein synthesis. Small terpenes are volatile, vaporizing easily, and so make up many of the essential oils associated with plants. The distinctive smells of lemon peel, mint, sage, menthol, pine resins, and geranium leaves are all due to terpenes. While we find these odors pleasant, they are feeding deterrents to mammals such as squirrels and moose. In addition, terpenes obstruct the growth and metabolism of both fungi and insects. As a result, pyrethrin, a terpene derivative extracted from chrysanthemums, is marketed commercially as an insecticide.
Other compounds derived from terpenes interfere with insect development. For example, phytoecdysteroids are plant-produced replicas of the hormones that induce molting in insects. Exposure to these compounds cause insects to molt prematurely and therefore keeps their populations in check. Taxadiene, precursor for the chemotherapy drug taxol, is a terpene that was first extracted from the bark of the Pacific yew tree.
Phenols form the third main class of defensive compounds, illustrated by the tannins found widely in plant tissues. Tannins bind with proteins, reducing their digestibility. Plants store tannins in cell vacuoles, and so these compounds come into contact with the protein-rich cytoplasm only when cells are damaged. Herbivores attempting to feed on tannin-producing plants obtain a poor reward for their efforts. For this reason, natural selection favors individuals that avoid tannin-rich plants. Many unripe fruits are high in tannins, and the unpleasant experience of biting into an unripe banana illustrates how tannins deter consumers. For thousands of years, humans have taken advantage of the protein-binding properties of tannins, using extracts from tree bark to process animal skins by “tanning” to produce leather.
Some of the chemical defenses used by plants are stored in a separate compartment from the enzymes that activate them. When you bite into a plant belonging to the cabbage family, the chemicals stored in the vacuole comingle with enzymes in the cytosol. The enzymes then catalyze the production of mustard oils from these compounds. Mustard oils give cabbage and its relatives their distinctive smell. They serve as feeding deterrents because they interfere with insect growth.
Cassava roots, an important food crop in Africa and South America, release the toxin hydrogen cyanide when their cells are damaged. Before cassava roots can be safely consumed, the roots must be ground and the cyanide-producing chemicals removed in running water. Although some cassava varieties, so-called sweet cassava, lack these chemicals and can be eaten without any special preparation, the bitter varieties are preferred crops in many cases because they often have higher yields.
A final chemical defense found in plants is protein based. Plants and animals use the same 20 amino acids to construct proteins, but some plants produce additional amino acids as well. Plants do not incorporate these additional amino acids into their proteins, but herbivores that ingest them do. The resulting proteins can no longer fulfill their function. Thus, insect herbivores that consume nonprotein amino acids grow slowly and often die early. A second protein-based defense is the production of antidigestive proteins called protease inhibitors. These proteins bind to the active site of enzymes that break down proteins in the herbivore’s digestive system. This prevents proteins from being broken down into their individual amino acids and therefore reduces the nutritional value of the plant tissue. Insects that feed on plants that produce protease inhibitors have reduced growth rates.
32.2.3 Some plants provide food and shelter for ants, which actively defend them.


While many plants have mechanical defenses against herbivores and nearly all have at least one form of chemical defense, a smaller number of species have evolved ecological defenses against herbivory. These plants “employ” animals as bodyguards in exchange for shelter or nourishment, much the way that many plants provide food or other rewards to animals that transfer pollen and disperse seeds (Chapter 30).
Nectar is typically associated with flowers. However, many plants produce nectar in glands located on their leaves (Fig. 32.12). These extrafloral nectaries, or nectar-producing regions, attract ants, which move actively throughout the plant in search of nectar. When the ants encounter either the eggs or larvae of other insects, they consume these as well. The value of this relationship to the plant can be demonstrated easily: When ants are prevented experimentally from patrolling certain branches, those branches suffer higher rates of herbivore damage.
A small number of plants, several hundred in total, have evolved a much closer relationship with ants. These so-called ant-plants provide both food and shelter for an entire colony of ants, which then defend their host. The bullhorn acacia (Acacia cornigera), which grows in dry areas of Mexico and Mesoamerica, produces hollow spines at the base of each leaf, as well as protein- and lipid-rich food bodies on the tips of its expanding leaves. Symbiotic ants (Pseudomyrmex ferruginea) live in the spines and feed on the food bodies (Fig. 32.13). To protect this source of food and shelter, the ants actively defend the plant. When the ants detect an invader, they produce an alarm pheromone that causes the entire colony to swarm over the plant, attacking any insects or vertebrates (including scientists) that they encounter. The ants will attack the growing tip of a vine that might climb up the host and even destroy plants growing in the soil beneath the host Acacia.
This system works well until it comes time to reproduce. Without some mechanism to control the hypervigilant ants, any pollinator that tried to visit one of the host plant’s flowers would be subject to the same treatment as a truly unwanted guest. To prevent pollinators from being turned away, flowers of the bullhorn acacia release a chemical that repels the resident ants, allowing pollinators to visit the flowers unharmed.
The interactions between ant-plants and their ants are highly coevolved. The ant species are found only in association with their host plants. And while ant-plants can be grown without ants, an undefended plant would rapidly be consumed in nature because ant-plants lack the chemical defenses present in related species that do not host ants.
32.2.4 Grasses can regrow quickly following grazing by mammals.
By far, the most important vertebrate herbivores are the bison, zebra, antelope, and their relatives that populate the great plains of East Africa and North America. Studies of herbivore damage in an East African savanna indicate that vertebrate herbivores annually consume approximately the same amount of leaf biomass as do herbivorous insects.
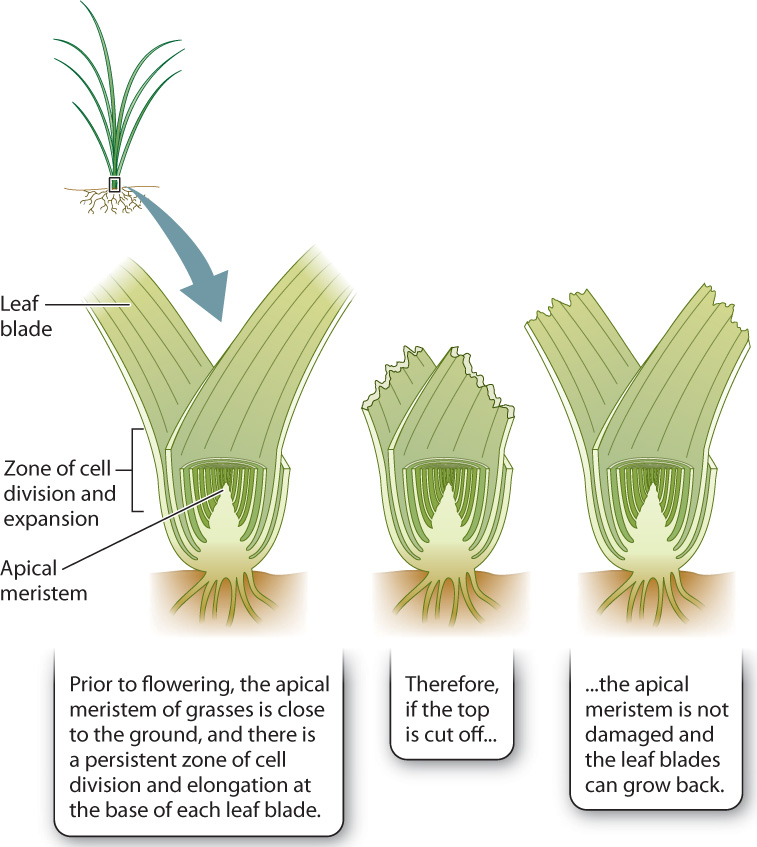
Grasses are well suited to cope with grazing mammals (Fig. 32.14). The vertically oriented leaves grow from stems with limited internodal elongation. This allows the shoot apical meristem to remain close to the ground, where it is less likely to be damaged. Grasses have persistent zones of cell division and elongation at the base of each leaf. Because grass leaves elongate from their base, grasses can replace leaf tissues removed by a grazing mammal, fire, or a lawn mower without having to add to the length of the stem. Only when grasses produce their flowering stalks does the apical meristem emerge above the leaves.
Grasses are thought to have evolved in forest environments. In fact, grass-dominated ecosystems have become widespread only during the past 25 million years. The fossil record shows that as grasslands expanded, grazing mammals evolved along with them. Horses, for example, originated as small browsers that fed on the leaves of small trees and shrubs. As grasslands expanded, larger horse species with teeth adapted to eating grass evolved. Why would specialized teeth be advantageous? The small plates of silica in grass cells appear to contribute to mechanical support and may provide mechanical defense against herbivores. Horses and other grazers that feed on these grasses wear down their tooth enamel grinding against the silica, so teeth with a thick enamel layer are favored by natural selection. It is not clear whether grasses evolved in response to grazing or to disturbances such as fire, but without question, their distinctive pattern of meristem activity minimizes the damage imparted by grazing mammals.