32.4 DEFENSE AND PLANT DIVERSITY
Many plants allocate a significant fraction of their resource and energy budgets to defensive chemicals and structures. Despite this commitment, insects consume approximately 20% of all plant production each year, while pathogens destroy a substantial additional amount. The magnitude of this loss means that pathogens and herbivores must be a potent force in both ecology and evolution, shaping competitive interactions by allowing some species to prosper and limiting the success of others. In nature, plant fitness may be strongly influenced by the capacity to deter herbivores and resist pathogens.
Interactions between plants and their consumers are thought to have influenced patterns of plant diversification. Just as pollinating insects diversified along with angiosperms (the flowering plants), so, too, did plant-eating insects. The fact that angiosperms and plant-eating insects radiated at the same time is consistent with an important role for plant–insect coevolution in generating diversity. Further evidence that defense has influenced plant diversification comes from plant genes and secondary compounds. For example, the R genes that are instrumental in pathogen recognition form one of the largest gene families in plants, while chemical defenses against herbivores include about 6000 alkaloids and more than 10,000 terpenoid compounds. Such observations suggest that plants are locked into an evolutionary arms race with herbivores and pathogens. This arms race is very much in evidence in the efforts by farmers to limit crop losses by reducing the numbers of the pathogens and herbivores in fields and orchards.
32.4.1 Pathogens, herbivores, and seed predators can increase plant diversity.
The lowland rain forests of eastern Ecuador have the highest diversity of tree species recorded anywhere: More than 1000 species of trees were identified in a 500 m × 500 m plot. What allows so many tree species to coexist is one of the great questions of modern biology. In the 1970s, two ecologists, Daniel Janzen and Joseph Connell, independently proposed that interactions with host-specific pathogens, herbivores, or seed predators play an important role.
The Janzen–Connell hypothesis, as it is now known, is based on the observation that seeds or seedlings often suffer high rates of mortality when growing directly beneath an adult tree of the same species (Fig. 32.18). Seedlings that grow farther away have a better chance of escaping pathogens that are common in the soil underneath an adult tree. By a similar argument, seeds that disperse far from their maternal plant may escape notice by seed predators, which often gather in higher densities near trees with mature fruit. Such density-dependent mortality provides rare species with an advantage over more common species. In other words, the offspring of a rare species is more likely to survive because its seeds are more likely to be dispersed into a location where there are no neighboring adult plants of the same species. By favoring the survival of less-common species, density-dependent mortality prevents one species from outcompeting all others.
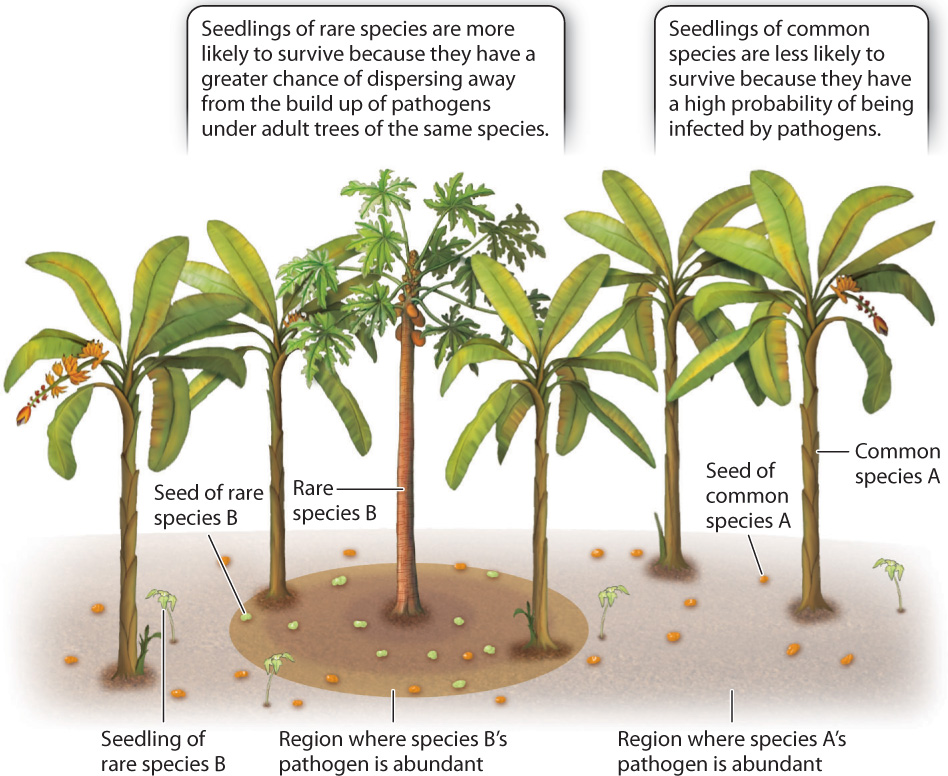
In the tropical rain forests of Ecuador and elsewhere, only a small number of tree species are common; the majority of tree species are rare. The ability to persist at low population density allows rare species to escape herbivore and pathogen damage, but requires reliable ways of getting pollen from one plant to another. Thus, animal pollination, herbivory, and infection by pathogens are thought to play complementary roles in promoting angiosperm diversity.
32.4.2 The evolution of new defenses may allow plants to diversify.
American scientists Paul Ehrlich and Peter Raven observed that closely related species of plants were fed upon by closely related species of butterflies. In 1964, they cited this observation as evidence that plants and their herbivores are caught up in a coevolutionary arms race: Plants evolve new forms of defense, and herbivores evolve mechanisms to overcome these defenses. In particular, Ehrlich and Raven hypothesized that the evolution of novel defenses has been an important force in the diversification of both plants and herbivorous insects.
An innovation in defense may allow a plant population to expand into new areas, increasing its potential to evolve into new species. Similarly, innovation in overcoming plant defenses may allow an insect or pathogen population access to new plant resources. Ehrlich and Raven’s “escape and radiate” hypothesis thus predicts a burst of diversification following the evolution of a key innovation that allows plants to avoid being eaten. This hypothesis makes intuitive sense, but only with the advent of phylogenetic trees based on DNA sequence comparisons (Chapter 23) could it be tested. Biologists knew that latex or resin canals evolved independently in more than 40 different groups of plants. Reasoning that these features represent a novel defense, they asked whether the lineages that evolved latex or resin canals were more diverse than closely related groups that lacked this innovation. In 14 of the 16 groups examined, the lineages with the protective canals were significantly more diverse than their closest relatives that lacked this innovation. This pattern supports the hypothesis that the evolution of latex and resin canals provided plants with the freedom to expand into new habitats.
Escalation of defenses, however, is not the only possible evolutionary outcome. Phylogenetic research shows that, as milkweeds diversified, many of the newly evolved species produced fewer chemical defenses such as latex canals and cardenolides and instead replaced tissues by growing quickly following damage. The presence of specialized herbivores such as monarch caterpillars that can disarm milkweeds’ defenses may help explain why these more recent plant species reduce investments in defense in favor of more rapid growth.
Case 6 Agriculture: Feeding a Growing Population
32.4.3 Can modifying plants genetically protect crops from herbivores and pathogens?
The density-dependence of herbivore and pathogen damage is a particular challenge for agriculture as practiced in most countries. When acre after acre are covered by a single crop species, successful invaders will not be slowed by low target density. Large expanses of a single crop allow pest populations to build up to high levels, and the tendency to plant genetically uniform varieties increases the losses in yield. Not surprisingly, then, herbivores and pathogens exact a major toll on agricultural crops, and protecting crops from damage is a high priority for farmers.
The production of pesticides and herbicides increased dramatically in the second half of the twentieth century. Together with irrigation and fertilizers, these compounds are important contributors to the increased yields of the Green Revolution. However, large-scale application of pesticides is not without risk. In addition to the potential for toxic effects, widespread chemical treatments provide a strong selective force for the evolution of resistance. As pests become resistant to pesticides, one response is to apply more or stronger chemicals. The consequences of this approach, however, can spiral out of control as chemical applications indiscriminately kill off the natural predators of the pests themselves.
During the 1980s, farmers applied increasing amounts of pesticides to rice paddies in South Asia, yet crop losses continued to increase. This dangerous course was reversed by the introduction of a program first developed in the 1950s, in which farmers devote substantial time and effort determining what pests are actually present, whether or not they are at damaging levels, and whether or not there are natural predators of the pests. Only after understanding the ecology of the farm are pests removed, starting with mechanical means, then biological control, and, only as a last resort, industrial pesticides. This approach, called integrated pest management, allows farmers to minimize routine pesticide use and therefore prevent pest species from evolving resistance.
To further improve the ability of crops to defend themselves against herbivores and pathogens, crop breeders must emulate—on a faster timescale—the role played by evolution in selecting for resistance to newly evolving pests. For example, Norman Borlaug’s original task was to develop wheat varieties for Mexico that were resistant to fungal pathogens. To do this, he obtained seeds of wheat varieties from around the world that showed evidence of pathogen resistance. He then crossed these disease-resistant varieties with ones adapted to the local conditions. Although it took thousands of hand-pollinations, Borlaug was successful at introducing resistance genes into wheat varieties that grew well in Mexico.
Crop breeding is a powerful way to alter the genetic makeup of cultivated species. However, because it relies on sexual reproduction, crop breeding is limited to the genetic diversity that exists within varieties that can interbreed. In contrast, Rhizobium radiobacter allows scientists to introduce specific genes found in distantly related organisms into cultivated species. One of the first commercial uses of this technology was to introduce chemical defenses from a bacterium into crop plants.
Bacillus thuringiensis (Bt), a soil-dwelling bacterium that produces proteins toxic to insects, has been used to control insect outbreaks in agriculture since the 1920s. At first, spores containing the toxins or the toxins themselves were applied directly to plants. In 1985, the genes that encode the toxins from B. thuringiensis were inserted into tobacco, where their expression conferred substantial protection against insects. Today, Bt-modified crops are some of the most widely planted genetically modified plants.

The introduction of Bt crops markedly reduced the application of pesticides, to the benefit of both farmworkers and the environment. However, the danger of planting Bt-expressing crops widely is that constant exposure to Bt toxins increases the probability that pest populations will evolve resistance. This possibility is of particular concern to organic farmers, who continue to use traditional application of B. thuringiensis spores to control insect outbreaks. To prevent the evolution of resistance, U.S. farmers are required by law to plant a fraction of their fields with non-Bt-expressing plants (Fig. 32.19). The proximity of non-Bt-expressing plants allows pests that are not resistant to Bt toxins to survive and interbreed with individuals feeding on the Bt-expressing plants, thus slowing the evolution of Bt-resistance. The efficacy of this practice, however, is called into question by reports of Bt-resistant pests and the need to apply increasing amounts of industrial pesticides to Bt crops.
If there is one practical lesson to be learned from our experience with Bt, it is that pathogens and herbivores will continue to evolve new ways of circumventing the defenses of crop plants. Agriculture will continue to require new forms of crop protection. While the use of chemical pesticides will probably remain widespread, crop protection will increasingly be based on the application of genetics, through both crop breeding and biotechnology. In either case, success will depend upon the availability of existing genes honed by natural selection. Without access to genes involved in pathogen recognition (R genes) or the production of chemical and mechanical defenses, we will have the tools, but not the raw materials, to protect our crops.
In some cases, crop protection will rely on genes from distantly related organisms, as illustrated by Bt. Commonly, however, defense strategies for crops will rely on genes from other plants, often ones that are closely related. For example, the specifi c R genes used for pathogen recognition typically function only within a single species or closely related species. Thus, recent eff orts to develop potatoes that are resistant to Phytophthora infestans focus on transferring R genes from a wild relative of potato. The continuation of such eff orts requires a commitment to safeguarding the genetic diversity found in natural populations of crop species and their close relatives. Only by tapping into these natural genetic resources can breeders produce crops that stay one step ahead of the constantly evolving threats of pathogens and herbivores.