33.4 GYMNOSPERMS
The seed—and equally important, pollen—are evolutionary innovations that played important roles in the success of seed plants. Seed plants do not require external water for fertilization. Instead, the male gametophyte is transported through the air in a pollen grain. Successful pollination leads to fertilization and the formation of a seed. Seeds are multicellular structures that enhance dispersal success by transporting the embryo together with resources that it can draw on during germination. Before these innovations, the ancestors of the seed plants evolved the ability to grow large woody stems through the formation of both a vascular and a cork cambium. Therefore, woody stems and roots are the ancestral condition in seed plants.
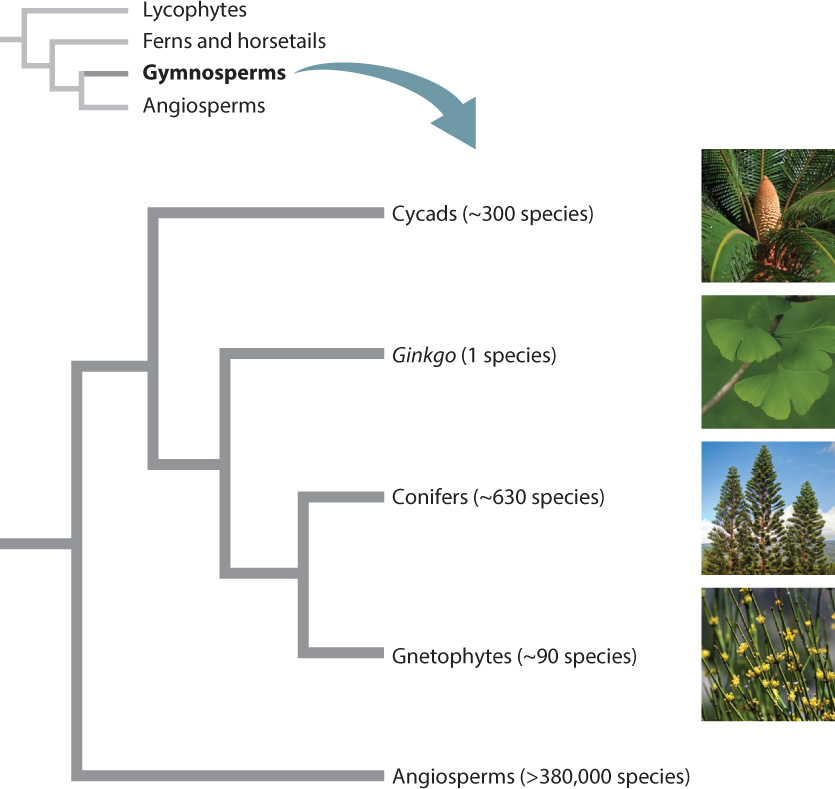
The gymnosperms are one of two major groups of seed plants. The living gymnosperms include four groups: cycads, ginkgos, conifers, and gnetophytes (Fig. 33.15). However, the fossil record shows that more than a dozen additional groups, now extinct, were once present. These include a number of early seed plants that branched off before any living gymnosperm groups evolved. The floodplains adjacent to the coal swamps in which giant lycophytes thrived were populated by early seed plants with large fern-like leaves that produced large fleshy seeds. Gymnosperms probably reached their greatest diversity during the time of the dinosaurs. At that time, groups like cycads and conifers lived alongside a larger number of now-extinct groups.
33.4.1 Cycads and ginkgos are the earliest diverging groups of living gymnosperms.
The unobservant eye often confuses cycads with palm trees because both have unbranched stems and large leaves (Fig. 33.16). However, the presence of large cones rather than flowers clearly marks cycads as gymnosperms. Cycads tend to have low photosynthetic rates and be slow growing. Although they have large stems, their vascular cambium produces little additional xylem, and most of their bulk is made up of a large pith and cortex. They form symbiotic associations with nitrogen-fixing cyanobacteria that allow them to grow in nutrient-poor environments. The apical meristem of cycads is protected by a dense layer of bud scales, enabling them to survive wildfires.

The approximately 300 species of cycads occur most commonly in tropical and subtropical regions. Most species have a limited and often fragmented distribution, suggesting that they were much more widespread in the past. Most cycad populations are small and often vulnerable to extinction. Cycads may owe their persistence in part to the fact that they rely on insects for pollination, as well as to their ability to live in nutrient-poor and fire-prone environments. Insects provide more efficient pollen transfer than wind, thus allowing species with small populations to reproduce successfully. Many cycads are pollinated by beetles, which are attracted by chemical signals produced by the cones. The insects use the large cones for shelter, and the pollen cones provide them with food. Following fertilization, brightly colored, fleshy seed coats attract a variety of birds and mammals that serve as dispersal agents. Because of their small numbers and fragmented distribution, two-thirds of all cycads are on the International Union for the Conservation of Nature’s “red list” of threatened species, the highest percentage of any group of plants.
Cycads are an ancient lineage, with fossils at least 280 million years old. During the time of the dinosaurs, they were among the most common plants. Cycad diversity and abundance declined markedly during the Cretaceous Period, at the same time as the angiosperms rose to ecological prominence. A recent study shows, however, that all the present-day species within this group date from a burst of speciation that took place approximately 12 million years ago. At that time, changing continental positions and ocean circulation patterns resulted in increasingly cooler and more seasonal climates, providing new habitats for emerging species while dooming older species to extinction.
A second group of gymnosperms has only a single living representative: Ginkgo biloba. Like cycads, ginkgos date back about 270 million years, and during the time of the dinosaurs, they were common trees in temperate forests. Also like cycads, ginkgos declined in abundance, diversity, and geographic distribution over the past 100 million years, as a result of both changing climates and angiosperm diversification. Ginkgo biloba forms tall, branched trees, with fan-shaped leaves that turn a brilliant yellow in autumn (Fig. 33.17). Once again, like cycads, Ginkgo develops fleshy seeds, perhaps a feature inherited from their common seed plant ancestor.

Ginkgos have long been cultivated in China, especially near temples, and it is unclear whether any truly natural populations exist in the wild. Today, however, Ginkgo once again has a global distribution, as its ability to tolerate pollution and other stresses associated with urban environments has made it a tree of choice for street plantings. The only drawback is that the fleshy seeds produce a chemical that smells like rancid butter. For this reason, individuals that produce pollen, as opposed to seeds, are strongly preferred.
33.4.2 Conifers are forest giants that thrive in dry and cold climates.
The approximately 630 species of conifers are, for the most part, trees. Conifers include the tallest (over 100 m) and oldest (more than 5000 years) trees on Earth (Fig. 33.18). Well-known conifers include pines, junipers, and redwoods. Conifer xylem consists almost entirely of tracheids, resulting in wood that is strong for its weight and has relatively uniform mechanical properties. Conifers have traditionally served as masts for sailing ships, and today are used for telephone poles. Conifers supply much of the world’s timber, as well as the raw material for producing paper.

Most conifers are evergreen, meaning that they retain their often needle-like leaves throughout the year. In some species, the leaves are remarkably long lived, remaining green and photosynthetic for decades. Many conifers produce resin canals in their wood, bark, and leaves that deter insects and fungi. Conifer resins are harvested and distilled to produce turpentine and other solvents. In addition, conifer resins are the source of a number of important chemicals, including the drug taxol, which is used in the treatment of cancer.
In Chapter 30, we examined the life cycle of pine. Pollen is produced in small cones, and ovules develop in larger cones that mature slowly as the fertilized ovules develop into seeds. Conifers are wind pollinated, and most species also rely on wind for seed dispersal. Some conifers, however, produce fleshy and often brightly colored tissues associated with their seeds that attract birds and other animals. In junipers, for example, the entire seed cone becomes fleshy. Juniper seeds are used as a seasoning; they are what gives gin its distinctive flavor.
Conifers dominate the vast boreal forests of Canada, Alaska, Siberia, and northern Europe. Conifers generally increase in abundance as elevation increases, and they are common in dry areas, such as the western parts of North America and much of Australia. Only a small number of conifer species are found in tropical latitudes, most commonly at higher elevations. Before the rise of the angiosperms, however, conifers were much more evenly distributed across the globe, including in the lowland tropics. One of the great questions in the evolutionary history of plants is why conifers were displaced from tropical latitudes by the angiosperms but continued to thrive in cold and dry environments.
An important difference between angiosperms and conifers is that angiosperms transport water in the multicellular xylem conduits called vessels, whereas conifers transport water in single-celled tracheids. In wet and warm environments, the xylem vessels of angiosperms can be both wide and long, increasing the efficiency of water transport through their stems. Conifer tracheids, which form from single cells, are much smaller. Thus, one hypothesis is that the evolution of highly conductive xylem vessels gave angiosperms a competitive advantage in many environments: They could support higher rates of photosynthesis and grow larger leaves. Once conifer populations began to dwindle, they could have disappeared completely because wind pollination, to be effective, requires large populations. In colder or drier regions, the dangers of cavitation due to drought or freeze followed by thaw (Chapter 29) constrain the size of angiosperm vessels. This helps level the playing field in the competition between conifers and angiosperms and explains the greater coexistence of conifers and angiosperms in cold or dry climates.
33.4.3 Gnetophytes are gymnosperms that have independently evolved xylem vessels and double fertilization.
Gnetophytes are a small group of gymnosperms (about 90 species) made up of three morphologically different genera (Fig. 33.19). Welwitschia mirabilis is found only in the deserts of southwestern Africa. It produces only two straplike leaves that elongate continuously from their base. Gnetum occur in tropical rain forests, where they form woody vines and small trees with large broad leaves. Ephedra, which is native to arid regions, produces shrubby plants with photosynthetic stems and small leaves.

Gnetophytes have long attracted the attention of plant biologists because their members exhibit a number of traits typically associated with angiosperms, notably the formation of multicellular xylem vessels and double fertilization, although the double fertilization in gnetophytes does not result in the formation of endosperm. Indeed, for years many botanists viewed gnetophytes as the sister group of angiosperms. DNA analysis, however, indicates that gnetophytes are not closely related to the angiosperms but are, instead, closely related to conifers. Thus, both vessels and double fertilization evolved independently and convergently in this group.
Quick Check 3
Double fertilization evolved independently in gnetophytes and flowering plants. Name two other features usually associated with angiosperms that evolved independently in gymnosperms.