36.1 ANIMAL SENSORY SYSTEMS
Animals are able to sense the physical properties of their environment, including light, chemicals, temperature, pressure, and sound, useful in finding mates and food and avoiding predators and noxious environments. Early in evolutionary history, organisms evolved specialized membrane receptors that functioned to detect these critical features in their environment. For example, before the evolution of nerve cells and a nervous system, bacteria evolved receptors located in the cell membrane to sense osmotic pressures that might otherwise rupture the membrane (Chapter 5). Bacteria and sponges also evolved membrane receptors that could detect chemicals—including nutrients—in their environment.
The senses of smell, taste, and sight in multicellular organisms with a nervous system also rely on membrane receptors. However, these receptors are embedded in specialized membranes of sensory neurons, forming a sensory receptor. In this case, a sensory receptor refers to the entire neuron, not just a membrane protein. In most multicellular animals, the sensory receptors are organized into specialized sensory organs that convert particular physical and chemical stimuli into nerve impulses that are processed by a nervous system and sent to a brain. Cnidarians (which include jellyfish, corals, and anemones) and roundworms evolved sensory receptors to sense physical contact, and cnidarians and flatworms were among the first multicellular animals to evolve simple light-sensing photoreceptive organs.
The conversion of physical or chemical stimuli into nerve impulses is called sensory transduction. For example, receptors located in the ear convert the energy of sound waves into nerve impulses that allow an animal to distinguish loud versus soft sounds and high-versus low-pitched sounds. Although the sense organs of different animals share many similar properties, differences have also evolved. Consequently, animals perceive the world differently from one another. Many insects, for instance, are sensitive to ultraviolet light; nocturnal snakes can see at night by sensing infrared radiation; and dogs can distinguish odor compounds as much as 100 million times lower in concentration than humans can detect (Fig. 36.1).

36.1.1 Specialized sensory receptors detect diverse stimuli.
Animals have evolved a diverse array of sensory receptors that respond to different stimuli, among them touch, light, and the oscillations in air pressure we call sound waves. How is a physical phenomenon in the world outside an organism transformed into a nerve impulse? The initial transformation takes place inside the sensory receptor (Fig. 36.2). A physical or chemical stimulus is converted into a change in the sensory receptor’s membrane potential. Recall from Chapter 35 that a cell’s membrane potential is the charge difference between the inside and the outside of the cell membrane, and that a depolarization in membrane potential is the first step in firing an action potential. Sensory receptors either fire action potentials themselves or synapse with other neurons that fire action potentials, which are then transmitted to the brain.
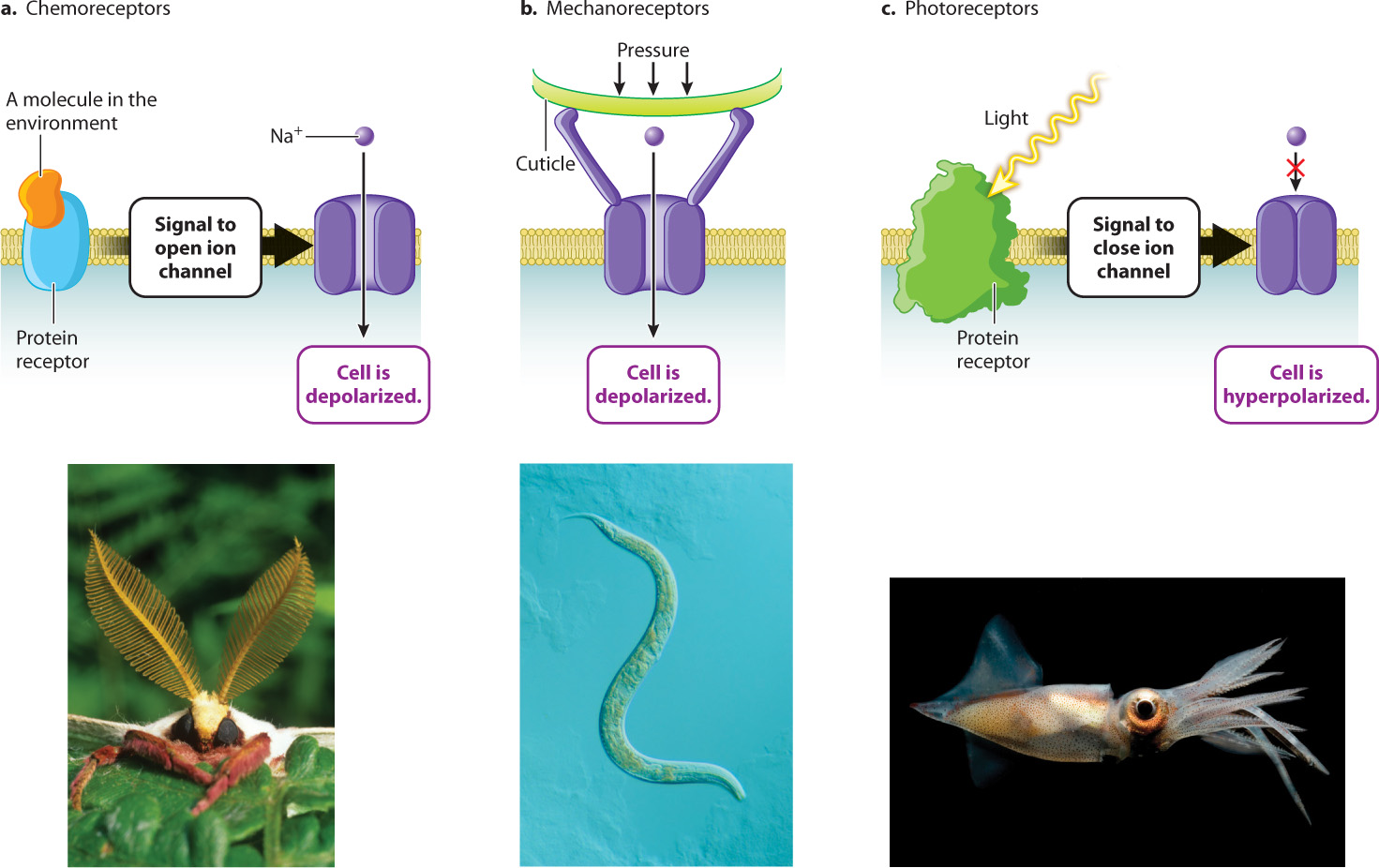
In many cases, the sound wave, touch, or other stimulus causes ion channels in the cell’s plasma membrane to open. The influx of ions changes the membrane potential by altering the distribution of charged ions on either side of the membrane. If the influx of ions reduces the charge difference, then the cell is depolarized (Chapter 35).
The most ancient type of sensory detection is chemoreception. All organisms respond to chemical cues in their environment. Even the earliest branching groups of Bacteria and Archaea have protein receptors in their cell membranes that respond to molecules in the environment. In animals, chemoreceptors respond to molecules that bind to specific protein receptors on the cell membrane of the sensory receptor (Fig. 36.2a). Many animals detect food in their environment by sensing key molecules such as oxygen (O2), carbon dioxide (CO2), glucose, and amino acids. Mosquitoes track CO2 levels to detect prey for blood meals, and coral polyps respond to simple amino acids in the water, extending their bodies and tentacles toward areas of greater concentration to feed. Other arthropods, such as flies and crabs, have chemosensory hairs on their feet. These animals taste potential food sources by walking on them. Salmon rely on chemoreception to detect chemical traces of the home waters of the river where they hatched, and where they will return to mate and spawn.
Chemoreception also underlies the sense of smell and taste. In most cases, the binding of molecules to a protein receptor on a taste receptor causes the protein receptor to change conformation. That change in conformation in turn triggers the opening of Na+ channels through G protein signal transduction pathways similar to those discussed in Chapter 9. The influx of Na+ ions depolarizes the receptor cell membrane. No action potential fires, but the depolarization travels far enough down the receptor’s short axon to trigger the release of neurotransmitters.
Mechanoreceptors respond to physical deformations of their membrane produced by touch, stretch, pressure, motion, and sound. Deformation of the receptor membrane opens sodium channels, causing a depolarization of the endings of the cell’s dendrites (Fig. 36.2b). Early mechanoreceptors in bacteria sensed local physical forces and internal cell pressure. Other mechanoreceptors in roundworms and anemones are linked to externally projecting cilia at the animal’s body surface.
One well-studied example of a mechanoreceptor is a touch receptor in the roundworm Caenorhabditis elegans. Deformation of the surface of the worm—its cuticle—exerts pressure on proteins connected to an ion channel. The mechanical force exerted changes the shape of the ion channel, causing it to open and leading to a change in membrane potential.
A mechanoreceptor in humans and other mammals is the sensory receptors found in the skin that sense touch and pressure. The cell bodies of these neurons are located in ganglia near the spinal cord, with extensions going in two directions: to the skin and to the spinal cord. In the skin, the neuron has branched tips, which are the initial sensors of touch and pressure. If the stimulus is strong enough, local depolarization leads to the firing of an action potential that travels all the way to the spinal cord. Thus, in contrast to a chemoreceptor, this type of mechanoreceptor transmits an action potential. The axon is much too long for simple depolarization to spread to its other end.
Mechanoreceptors are also found at the base of whiskers that rodents, cats, dogs, and other mammals use to sense touch with their snouts. Stretch receptors found in muscles are also mechanoreceptors that influence a muscle’s motor activation, helping to control its length and force. A very different group of specialized mechanoreceptors called hair cells are the sensory receptors for balance, gravity sensing, and hearing, discussed in section 36.3.
Thermoreceptors in the skin and in specialized regions of the central nervous system (CNS) respond to heat and cold. Thermoreceptors help to control an animal’s metabolism and patterns of blood flow, regulating body temperature by affecting rates of heat gain and loss (Chapter 40). As a result, they help to maintain homeostasis. Many invertebrate animals rely on thermoreception when seeking environments that provide favorable temperatures. Thermoreceptors in the skin of vertebrate animals are believed to be simple dendrite endings that respond to changes in skin temperature by altering the firing rate of the cell’s action potentials.
Nociceptors, or pain receptors, are another class of nerve cell with dendrites in the skin and connective tissues of the body. When these sensory receptors are exposed to an excessive mechanical, thermal, or chemical stimulus, they send action potentials to the brain or spinal cord, typically stimulating rapid withdrawal from the painful stimulus. Pain is a subjective sensation, and therefore difficult to assess and treat. Although it is clear that many animals experience pain, it is difficult to judge how different species associate the sensation of pain with underlying physical or chemical stimuli.
Light-detecting photoreceptors are the sensory receptors in eyes (Fig. 36.2c). Photoreceptors respond to individual photons of light energy by closing Na+ channels, causing the cell membrane to become hyperpolarized rather than depolarized. Most receptors excite the neurons that they synapse with, but vertebrate photoreceptors are unusual in that they inhibit the firing rate of other neurons within the eye rather than exciting them. Photoreception in the vertebrate eye is discussed in section 36.4.
Some fish, such as catfish, contain specialized electroreceptors arranged in a lateral line along their bodies. Electroreceptors enable these fish to detect weak electrical signals emitted by all organisms. They likely evolved as an adaptation for locating prey or potential predators in poorly lit habitats where vision was less useful. Some specialized “weakly electric” fish actually generate an electromagnetic field by emitting pulses from an electric organ located in the tail. Disturbances in the field detected by electroreceptors of the lateral line system signal the location of nearby prey. These fish also inhabit rivers with poor visibility. The bill of the duckbilled platypus also contains electroreceptors that locate prey in dimly lit water.
Quick Check 1
Aspirin and ibuprofen reduce pain by inhibiting the synthesis of chemicals called prostaglandins. What effect would you predict prostaglandins to have on the firing rate of nociceptors?
36.1.2 Stimuli are transmitted by changes in the firing rate of action potentials.
Sensory reception depends on converting the energy of a physical or chemical stimulus into an action potential, either in the sensory receptor itself or in the neuron it synapses with. How do these nerve impulses convey information to the brain? In effect, action potentials can be considered a code that the brain deciphers.
To convert information from the environment into a code, the nerve impulses conveyed by sensory organs carry out the following functions:
- Convey the strength of the incoming signal—the brightness of light, the loudness of a sound, or the intensity of an odor.
- Carry information about even weak signals when necessary.
- Convey the location of a signal’s source—for example, where is the body being touched?
- Filter out unimportant background signals.
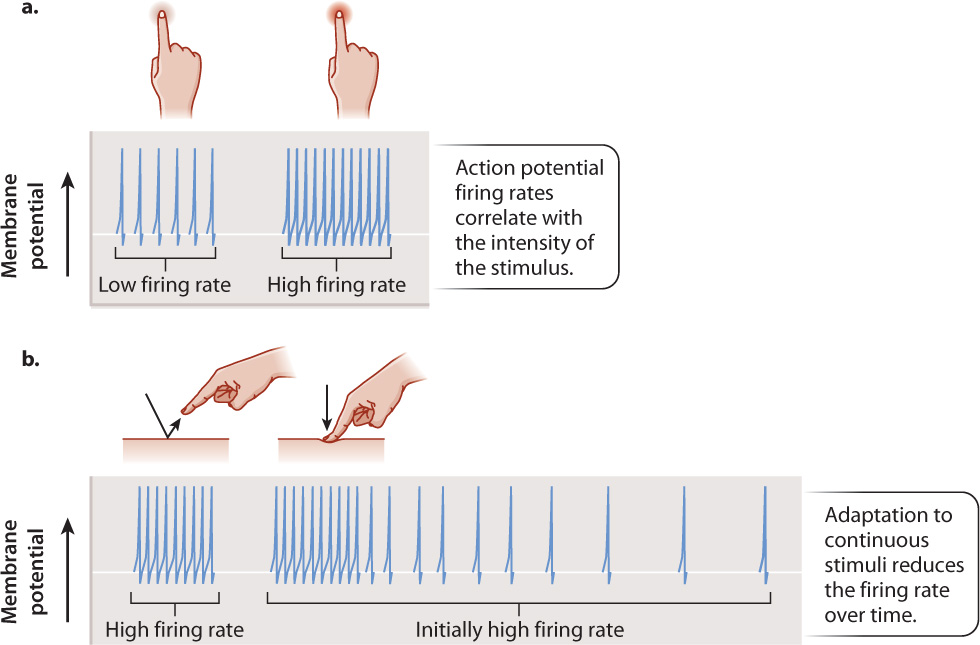
Because nerve cells transmit action potentials in an all-or-nothing fashion, information about the strength of the signal is coded by the number of action potentials fired over a period of time, which is the nerve cell’s firing rate. Most commonly, strong signals produce high firing rates and weak signals produce low or background firing rates (Fig. 36.3a). For example, the firing rate of mechanoreceptors increases as the tactile pressure they sense increases.
Several mechanisms increase a nerve cell’s sensitivity to even weak signals. Following sensory transduction, the signal typically undergoes initial processing. As discussed in Chapter 35, sensory stimuli may be summed over space or time. Multiple receptors that receive a stimulus often converge onto a neighboring neuron that increases its firing rate proportionally to the number of signals received. This is an example of spatial summation. Combining the synaptic input increases the receptor organ’s sensitivity to external stimuli. Temporal summation is the integration of sensory stimuli that are received repeatedly over time by the same sensory cell. When a sensory cell is stimulated more frequently, the excitatory postsynaptic potentials (EPSPs) sum, making it more likely that the cell is depolarized above threshold and fires an action potential.
Sensory receptors also distinguish between discrete (that is, intermittent) and continuous stimuli, providing a way to ignore background noise. Novel stimuli are generally the most important for an animal to respond to. Consequently, sensory receptors initially respond most strongly when a stimulus is first received (Fig. 36.3b). If the stimulus continues over a longer time period, sensory receptors typically reduce their firing rate through a process called adaptation (this is not the adaptation that results from natural selection). Adaptation of firing rate enables an animal’s sensory receptors to focus on novel stimuli, which are those likely to be most important. You can experience adaptation by touching your skin lightly at a particular location. Initially, skin mechanoreceptors fire in response to the touch, but after a while their firing rate diminishes and the sensation of a touch becomes less noticeable. Adaptation also ensures that the clothes touching your skin do not cause an irritating and distracting long-term sensation.
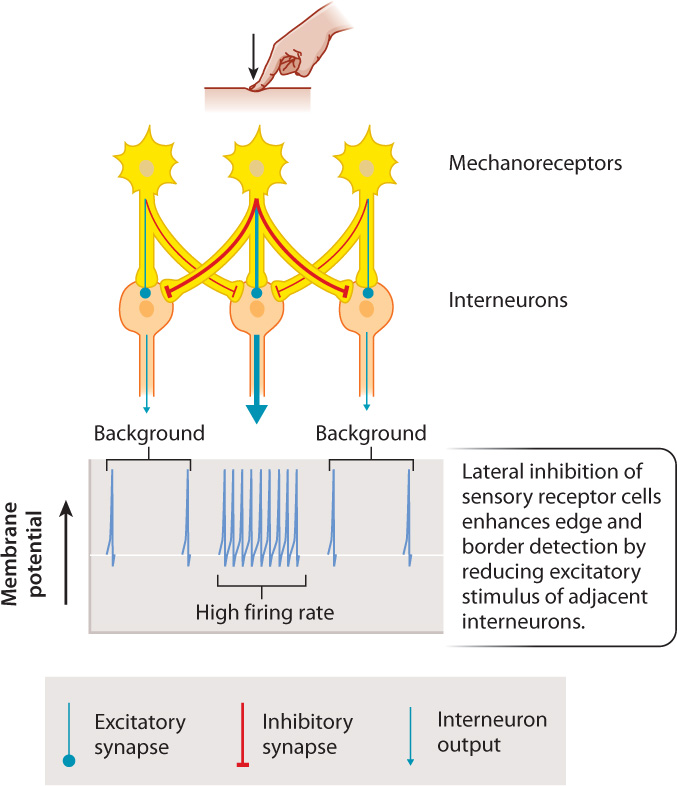
In addition, sensory receptors locate a signal’s source by lateral inhibition (Fig. 36.4). Lateral inhibition enhances the strength of a sensory signal locally but diminish it peripherally. In this process, receptors inhibit interneurons that receive sensory inputs from adjacent regions of the sensory organ, while sending excitatory signals to interneurons within their local region. This increases the contrast in signal strength between adjacent regions. For example, lateral inhibition in the retina of the eye enhances edge detection and sharpens image contrast. It also enables the sense of touch to be localized to a small region of the body surface. The inhibitory and excitatory signals are transmitted by synapses that have different neurotransmitters.
We have not yet described all the information in the code. How do action potentials convey pitch or color or a particular odor? This information is conveyed by the identity of the particular sensory receptors that are activated. Sensory organs are so constructed that, for example, tone A activates “tone A” sensory neurons in the ear, red light activates “red” sensory neurons in the eye, and the fragrance of a rose activates “rose” sensory neurons in the nose. The way in which the “correct” sensory neuron is excited varies for each of the senses. In the next three sections, we discuss how this happens for smell and taste, hearing, and vision.