37.2 MUSCLE CONTRACTILE PROPERTIES
Actin and myosin are evolutionarily conserved across all animal life, so all muscles have similar force-generating properties. The huge muscles of a weight lifter are evidence that larger muscles can exert more force. Why is that? The summed force produced by adjacent myofibrils of activated muscle fibers determines the force the whole muscle can exert. This means that larger muscles with more fibers produce greater forces than smaller muscles with fewer fibers. However, the force that a muscle produces also depends on its contraction length and the speed of contraction, as we discuss next.
37.2.1 Muscle length affects actin-myosin overlap and generation of force.
Why is it difficult to jump while standing on your toes? A muscle’s ability to generate force depends in part on how much it is stretched before contraction begins. The sliding filament model for muscle contraction, like any good model, makes specific predictions. In this case, it predicts that the amount of overlap between actin and myosin filaments within the sarcomere determines the number of cross-bridges that can form. The more overlap, the more force that can be produced. Conversely, when a muscle fiber is pulled to long lengths and then contracts, it generates lower force because the overlap between myosin and actin filaments is reduced. When a muscle fiber contracts at short lengths, it also produces lower force because myosin filaments begin to run into the Z disc at the end of the sarcomere, disrupting the geometry of the myosin filament lattice and hindering cross-bridge formation. At intermediate lengths the muscle fiber generates maximum force because actin–myosin filament overlap is greatest. These predictions were tested by experiments (Fig. 37.9).
FIG. 37.9How does filament overlap affect force generation in muscles?
BACKGROUND From the 1930s through the 1960s, British muscle physiologists studied the properties of striated frog muscle fibers. In the first set of experiments, A. V. Hill examined how a muscle’s shortening velocity affects its force production. In the second set of studies, British physiologists Andrew Gordon and Andrew Huxley, together with American physiologist Fred Julian, studied how a muscle’s contractile length affects its force-generating ability.
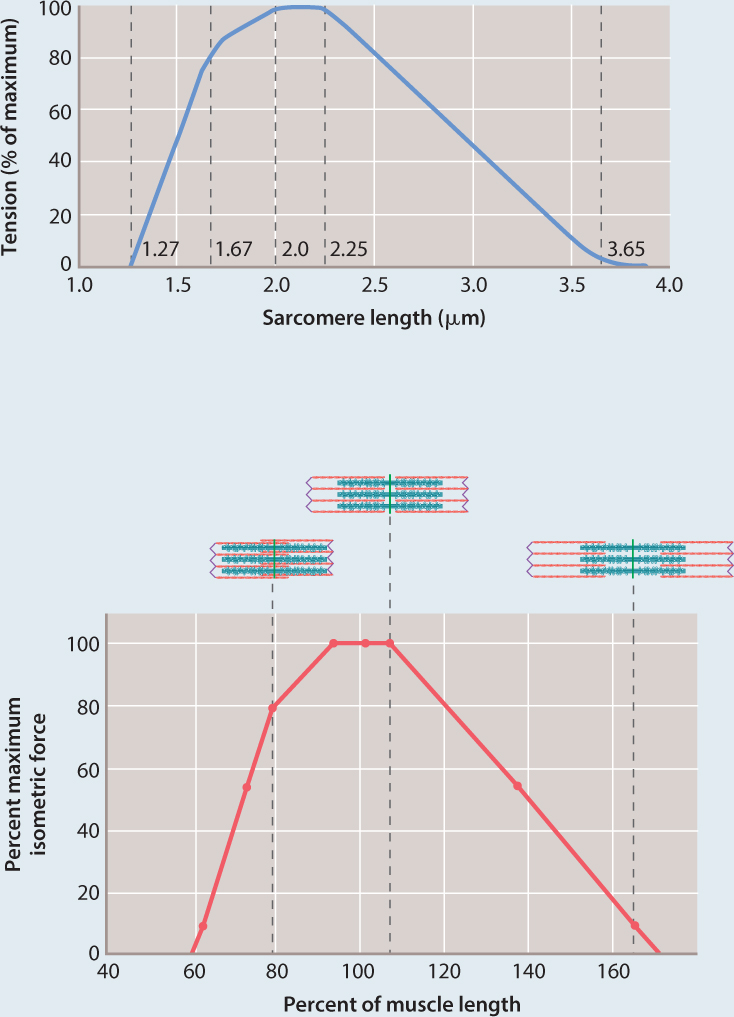
HYPOTHESIS They hypothesized that changes in overlap between myosin and actin affect the number of cross-bridges and thus the force the muscle can produce.
EXPERIMENT Gordon, Huxley, and Julian isolated a single muscle fiber and used optical microscopy to measure sarcomere length. They kept this length constant while the fiber was stimulated to produce force. Measurements of force were obtained at different sarcomere lengths.
RESULTS The top graph shows the data charted by Gordon, Huxley, and Julian, and the bottom graph reinterprets their data in terms of force and percent of muscle length. Their experiments showed that the muscle fiber produced maximal force at an intermediate sarcomere length (approximately 2.3 µm). At this length, the greatest number of cross-bridges were hypothesized to form, generating maximal force. As the sarcomere length was decreased or stretched, and then stimulated to contract, force declined. These results were consistent with a decrease in the number of cross-bridges that could form as a result of decreased actin–myosin overlap.
FOLLOW-UP WORK Other studies confirmed that changes in myosin cross-bridge formation are linked to changes in actin and myosin filament overlap at different sarcomere lengths, supporting the sliding filament model of muscle contraction.
SOURCE Gordon, A. M., A. F. Huxley, and F. J. Julian. 1966. “The Variation in Isometric Tension with Sarcomere Length in Vertebrate Muscle Fibres.” Journal of Physiology 184 :170–192.
This influence of muscle fiber length on force generated is the reason that you can’t jump well if you are on your toes and your ankles are fully extended: This position shortens your calf muscle fibers. Conversely, if your ankles are overly flexed, your calf muscles are lengthened, and again jumping force is limited. Through training, athletes and dancers learn techniques, such as bending the knees before jumping, so that their leg muscles contract at intermediate lengths to maximize muscle-force output.
The relationship between force and length of striated muscle also explains Starling’s Law for the function of the heart as a pump (discussed in Chapter 39), which ensures that the heart contracts more strongly when it is filled by larger amounts of blood returning from the veins.
Quick Check 3
Why can you lift a larger load when your elbow is slightly flexed and your biceps muscle is at an intermediate length than you can when your elbow is fully extended?
37.2.2 Muscle force and shortening velocity are inversely related.
In experiments carried out in the 1930s, A. V. Hill (who had won the 1922 Nobel Prize in Physiology or Medicine for his work on muscle energy use) observed that a muscle shortens fastest when producing low forces (Fig. 37.10). To produce larger forces, the muscle must shorten at progressively slower velocities. For example, your arm moves faster when you throw a light ball than when you throw a heavy one.
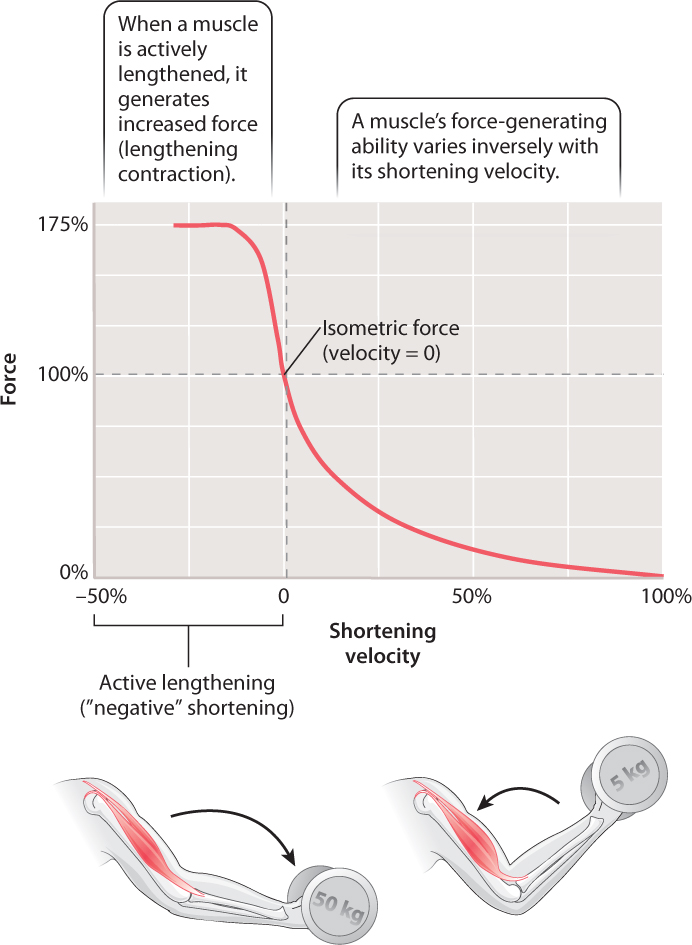
The term “muscle contraction” suggests that muscles shorten when stimulated to generate force (in a shortening contraction), but in fact muscles may also exert force and remain a uniform length or even be lengthened. Consequently, the term “contraction” needs to be broadened beyond its familiar reference to muscle shortening to include those instances when a muscle exerts force while remaining the same length or being actively stretched.
When a muscle contracts but cannot shorten (for example, when you carry a heavy bucket of water), the muscle’s shortening velocity is zero (Fig. 37.10), and it generates an isometric force (iso- means “same” and metric refers to “measure”). That is, the muscle still generates force, even though it stays the same length and does not shorten. Isometric exercises use this type of muscle contraction: Holding a weight in a fixed position or exerting force against an immovable object helps maintain muscle strength and may be beneficial in rehabilitation after injury by avoiding joint motion.
Muscles can also be stretched when the external load against which they contract exceeds their force output. This action is known as a lengthening contraction. During a lengthening contraction, the muscle generates much more force (up to 75% more, Fig. 37.10) than when it remains isometric or shortens. This kind of contraction occurs when you lower a box from a shelf or when your leg muscles are stretched to support your body in landing from a jump. It also happens during every stride you take when running. A brief phase of muscle stretch at the onset of contraction is common in animal movement because it allows the muscles to generate more force than if they remained isometric or if they began shortening at the start of the contraction. Muscle lengthening also reduces the muscle’s energy consumption because fewer fibers are recruited to generate a given level of force.
Injury can result if a muscle is too rapidly or repetitively stretched. Rapid and repetitive stretching often occurs during prolonged downhill hiking, causing muscle damage and soreness, particularly in the quadriceps muscles in the front of the thigh. Suitable training and warm-up exercises help muscles to stretch without damage.
37.2.3 Antagonist pairs of muscles produce reciprocal motions at a joint.
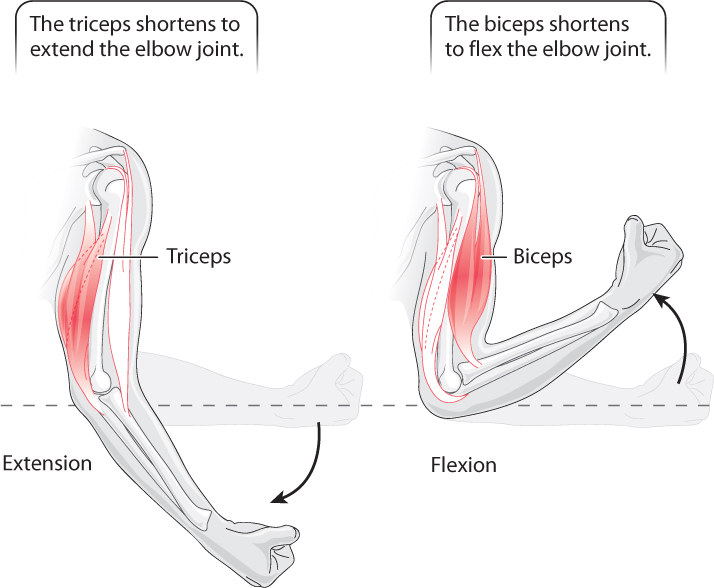
Because muscles can generate force only by pulling, not pushing, on the skeleton, to produce reciprocal joint and limb movements they must be arranged as pairs of antagonist muscles that pull in opposite directions. Flexion is the joint motion in which bone segments rotate closer together, and extension describes the motion when they move apart. For example, muscles at the elbow joint are organized as flexors (the biceps muscle) that contract to bring the arm toward the shoulder, and extensors (the triceps muscle) that contract to extend the arm (Fig. 37.11). Additional pairs of antagonist muscles must be present for movements in other directions. Muscles in the body wall of cnidarians (jellyfish and sea anemones) and segmented annelid worms, as well as the smooth muscles in the walls of intestines, are organized in longitudinal (lengthwise) and circumferential (circular) layers, allowing them to function as muscle antagonists that control movement and shape.
In contrast, when muscles combine to produce similar motions they are termed muscle agonists. Muscles arranged as agonists increase the strength and improve the control of joint motion. The three heads of the triceps are agonists of each other (Fig. 37.11).
37.2.4 Muscle force is summed by an increase in stimulation frequency and the recruitment of motor units.
How are animals able to vary the amount of force exerted by a muscle? As we have seen, skeletal muscles are stimulated by motor nerves, which conduct action potentials to the neuromuscular junction. The force exerted by a muscle depends on the frequency of stimulation by the motor nerve because more-frequent stimulation increases the amount of calcium that is released to activate cross-bridge formation in the muscle fibers. A single action potential results in a twitch contraction of a certain force (Fig. 37.12a). If sufficient time is allowed for the muscle to pump Ca2+ back into the sarcoplasmic reticulum, the contraction ends, force falls to zero, and a second stimulus can elicit a twitch contraction of the same force. However, if a second action potential arrives before the muscle has relaxed, a greater force is produced. With increasing frequency of stimulation, muscle force increases (Fig. 37.12b). This property of muscle contraction is called force summation.
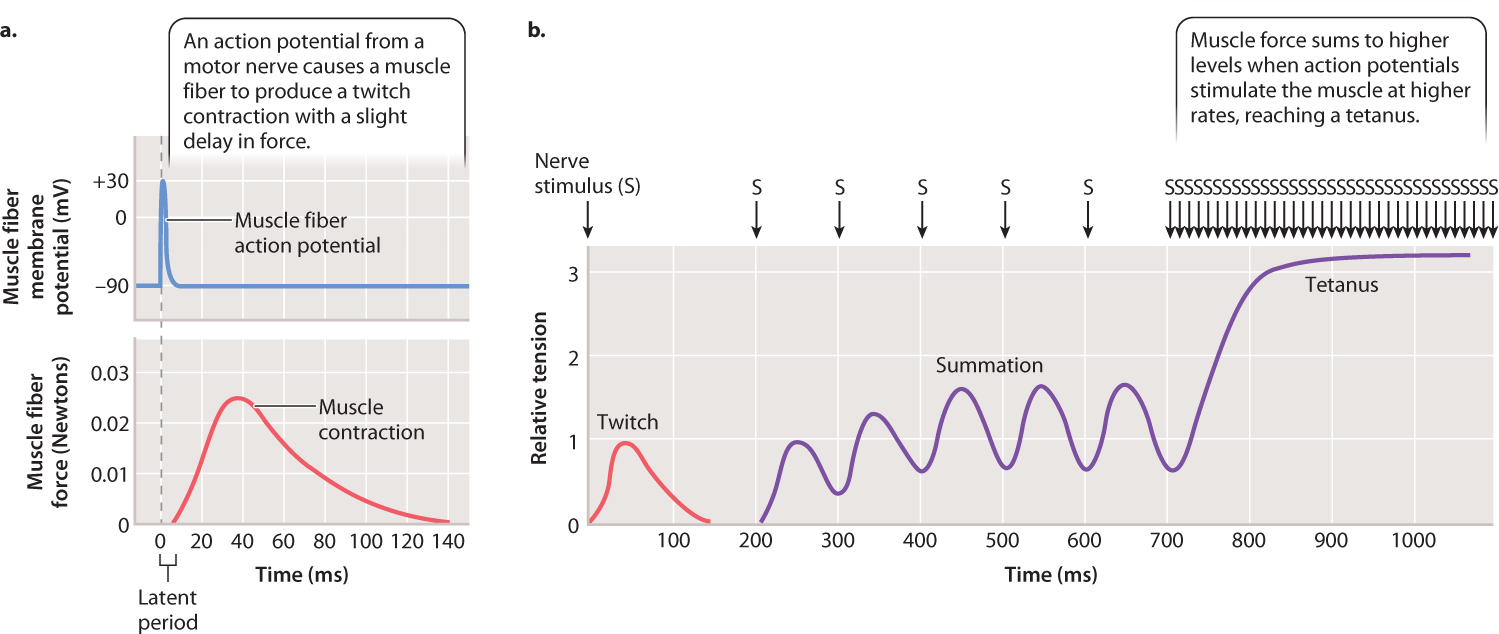
At a sufficiently high stimulation frequency, muscle force reaches a plateau and remains steady as stimulation continues. This muscle contraction of sustained force is described as a tetanus. Measurements in animals indicate that skeletal muscles commonly produce tetanic contractions, which generate steady and large forces. Tetanus is a normal physiological response of a muscle, but it can also be induced by the bacterium Clostridium tetani, so named because it causes severe muscle spasms, particularly of the jaw, chest, back, and abdomen.
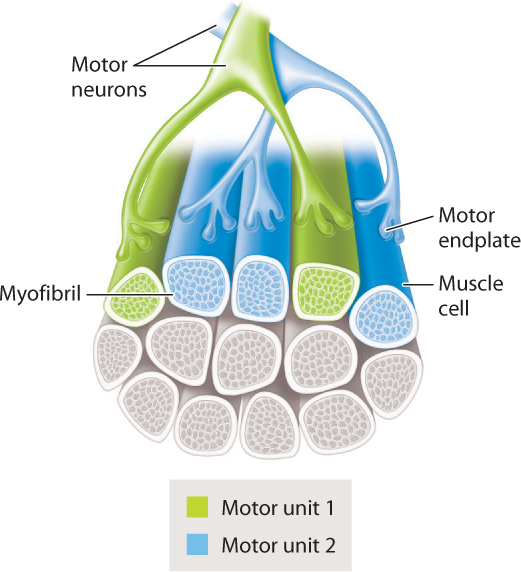
A motor neuron and the population of muscle fibers (cells) that it innervates is collectively termed a motor unit (Fig. 37.13). Motor units can include relatively few to several hundred muscle fibers. The number of muscle fibers innervated by a given motor nerve, and hence the size of the motor unit, affects how finely a muscle’s force can be controlled. For example, finger muscles have small motor units allowing fine adjustments in muscle force, whereas leg muscles have large motor units providing larger adjustments in force. Together with stimulation frequency, a muscle’s force output depends on the number of motor units (and therefore the number of muscle fibers) that are activated. Within each motor unit, muscle fibers generally share similar contractile and metabolic properties. Differences in these properties can give muscles very different capabilities, as discussed in the next section.
37.2.5 Skeletal muscles have slow-twitch and fast-twitch fibers.
The difference between a sprinter and a marathoner is not just training but also the properties of their muscles. There are different types of muscle fiber (Fig. 37.14a), allowing variation in the way muscles contract. In vertebrates, these are most simply classified as red slow-twitch fibers and white fast-twitch fibers. Slow-twitch fibers are found in muscles that contract slowly and consume less ATP to produce force. An animal uses slow-twitch fibers to control its posture or move slowly and economically. Fast-twitch fibers generate force quickly, producing rapid movements, but they consume more ATP. An animal depends on fast-twitch fibers to sprint or lunge at its prey. A key difference between the two types of fiber is that slow-twitch fibers obtain their energy through oxidative phosphorylation (aerobic respiration), and fast-twitch fibers obtain their energy mainly through glycolysis (Chapter 7).
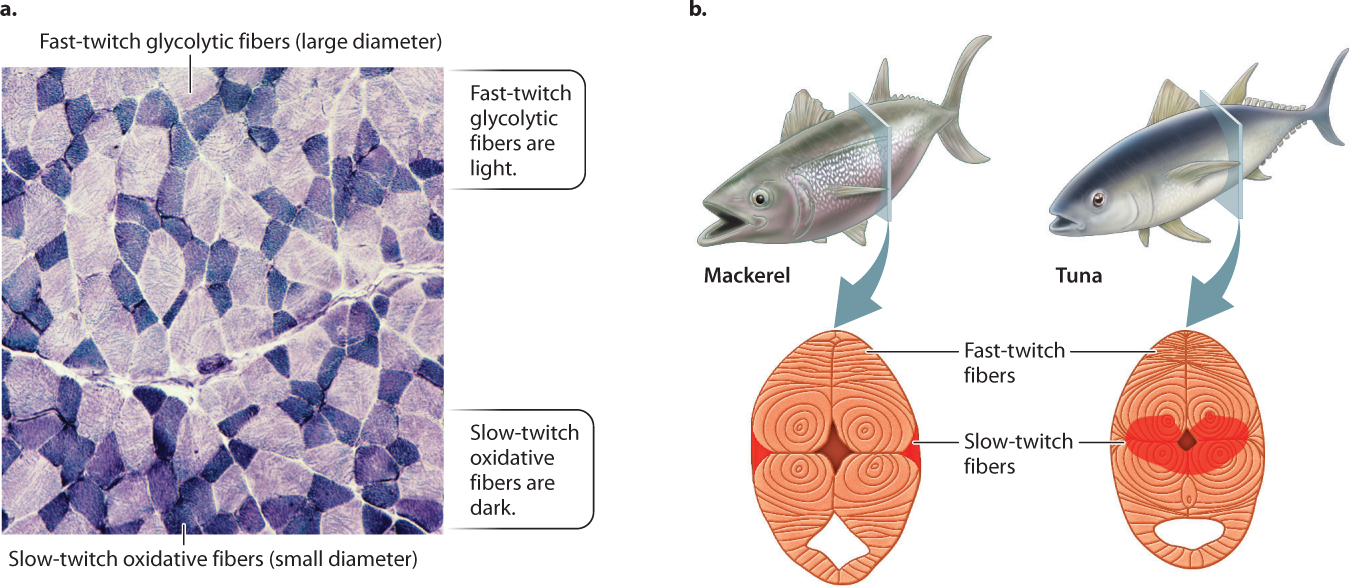
Although slow-twitch fibers develop force more slowly, they have greater resistance to fatigue in response to repetitive stimulation—that is, their loss of force over time is less. Their greater endurance is explained by the ability of their mitochondria to supply ATP to muscle fibers by means of aerobic respiration. Slow-twitch fibers have many mitochondria and are well supplied by capillaries. They contain an abundance of myoglobin, an oxygen-binding protein related to hemoglobin that facilitates oxygen delivery to the mitochondria (Chapter 39). The iron in myoglobin gives muscles with these fibers a red appearance. Slow-twitch muscle fibers express a chemically “slow” form of myosin with a relatively low rate of ATP hydrolysis, limiting their speed of cross-bridge cycling and force development.
White fast-twitch muscle fibers have fewer mitochondria and capillaries, contain little myoglobin, and rely heavily on anaerobic glycolysis (lactic acid fermentation) to produce ATP. Fast-twitch fibers express a chemically “fast” form of myosin with a high rate of ATP hydrolysis, favoring rapid force development and movement. However, fast-twitch fibers fatigue quickly because of the drop in pH caused by the accumulation of lactic acid. Because white fibers are larger than red fibers, they generate more force. In general, most skeletal muscles are composed of mixed populations of red and white fiber types, providing a broad range of contractile function.
Athletes who have more oxidative slow-twitch fibers excel at endurance and long-distance competitions. In contrast, athletes who have larger numbers of fast-twitch fibers perform better in sprint races and weight-lifting competitions. Because the distribution and properties of muscle fiber types are strongly inherited, it is largely true that champions are born and not made. Nevertheless, aerobic training can significantly increase the energy output through oxidative phosphorylation of an individual’s muscle fibers, just as weight lifting and speed training can enhance an individual’s strength and sprinting ability. Contrary to popular belief that weight lifting builds muscle by straining the muscle to build new fibers, it actually builds muscle by increasing the size of existing muscle fibers through the synthesis of additional myosin and actin filaments. This synthesis increases the cross-sectional area of the fiber (cell) and thus its force-generating capacity.
Case 7 Predator–Prey: A Game of Life and Death
37.2.6 How do different types of muscle fiber affect the speed of predators and prey?
We have just seen that slow-twitch muscle fibers provide endurance for sustained activity, whereas larger fast-twitch fibers produce greater speed and strength but fatigue quickly. The relative distribution of these fiber types in the muscles of different animals is influenced mainly by genetic heritage. This affects the locomotive behavior and capacities of both predator and prey animals. Whereas cheetahs have exceptional speed, their muscles tire quickly. As their prey, antelope have muscles that enable endurance long-distance running and contract rapidly for maneuvering.
Highly aerobic animals such as dogs and antelope have large fractions of slow-twitch fibers and specialized fast-twitch muscle fibers that also have a high oxidative capacity in addition to using glycolysis for rapid ATP synthesis. These muscle fibers allow the animals to move for extended periods of time at relatively fast speeds. In contrast, cats have mainly fast-twitch fibers, enabling them to sprint and pounce quickly. Nevertheless, cats and other animals use particular muscles with high concentrations of slow-twitch fibers (these are the dark-staining fibers shown in Fig. 37.14a) for slower postural movements and stealth. Animals such as sloths and lorises that move slowly have mostly oxidative slow-twitch fibers and few glycolytic fast-twitch fibers.
More generally, animals with high body temperatures and high metabolic rates (birds and mammals) have larger numbers of oxidative fibers compared with animals that have lower body temperatures and lower metabolic rates (reptiles). As a result, lizards use glycolytic fast-twitch fibers to sprint for brief periods but must then recover from the acid buildup produced by their muscles’ anaerobic activity in longer periods of rest (Chapter 40). In contrast, birds and mammals can sustain activity over longer time periods because they have oxidative muscle fibers supported by aerobic ATP supply.
Most fish have a narrow band of slow-twitch red muscle fibers that runs beneath their skin, which they use for slow steady swimming. Tuna and some sharks have evolved deeper regions of red muscle that they can keep warm (Fig. 37.14b), enabling their muscles to contract longer and faster to enhance their swimming. When escaping from a predator or attacking prey, fish recruit their larger fast-twitch white trunk musculature for rapid swimming.
The evolutionary arms race between predators and prey has resulted in selection for particular muscle fiber types and other musculoskeletal specializations for rapid movement and maneuvering. Features of the skeleton that contribute to these specializations are discussed next.