37.3 ANIMAL SKELETONS
Most animal skeletons provide a rigid set of elements that articulate, that is, they meet at joints, to transmit muscle forces for movement and body support. They also enable many animals to manipulate their environment by digging burrows, building nests, or constructing webs to catch food. Animals have evolved three types of skeletal system: hydrostatic skeletons, endoskeletons, and exoskeletons. All three types of skeleton use rigid elements to resist the pull of antagonist sets of muscles.
37.3.1 Hydrostatic skeletons support animals by muscles that act on a fluid-filled cavity.
Hydrostatic skeletons evolved early in multicellular animals with the first cnidarians (jellyfish and anemones) approximately 600 million years ago. They are found in nearly all multicellular animals as well as in many vascular plants. In animals that depend on a hydrostatic skeleton, fluid contained within a body cavity serves as the supportive component of the skeleton. Muscles exert pressure against the fluid to produce movement.
Two sets of muscles surround the fluid, controlling the width and length of the body cavity and, in many cases, of the whole animal (Fig. 37.15a). Circular muscles reduce the diameter of the body cavity, and longitudinal muscles reduce its length. A sea anemone can bend its body in various directions to feed or to resist water currents by closing its mouth to keep its fluid volume constant and contracting its circular and longitudinal muscles. Sea anemones extend themselves into the water column by contracting their circular muscles and relaxing their longitudinal muscles. Controlling the amount of fluid in their body cavity also allows sea anemones to adjust their shape. When threatened, a sea anemone rapidly retracts by contracting its longitudinal muscles and allowing water to escape from its body cavity.
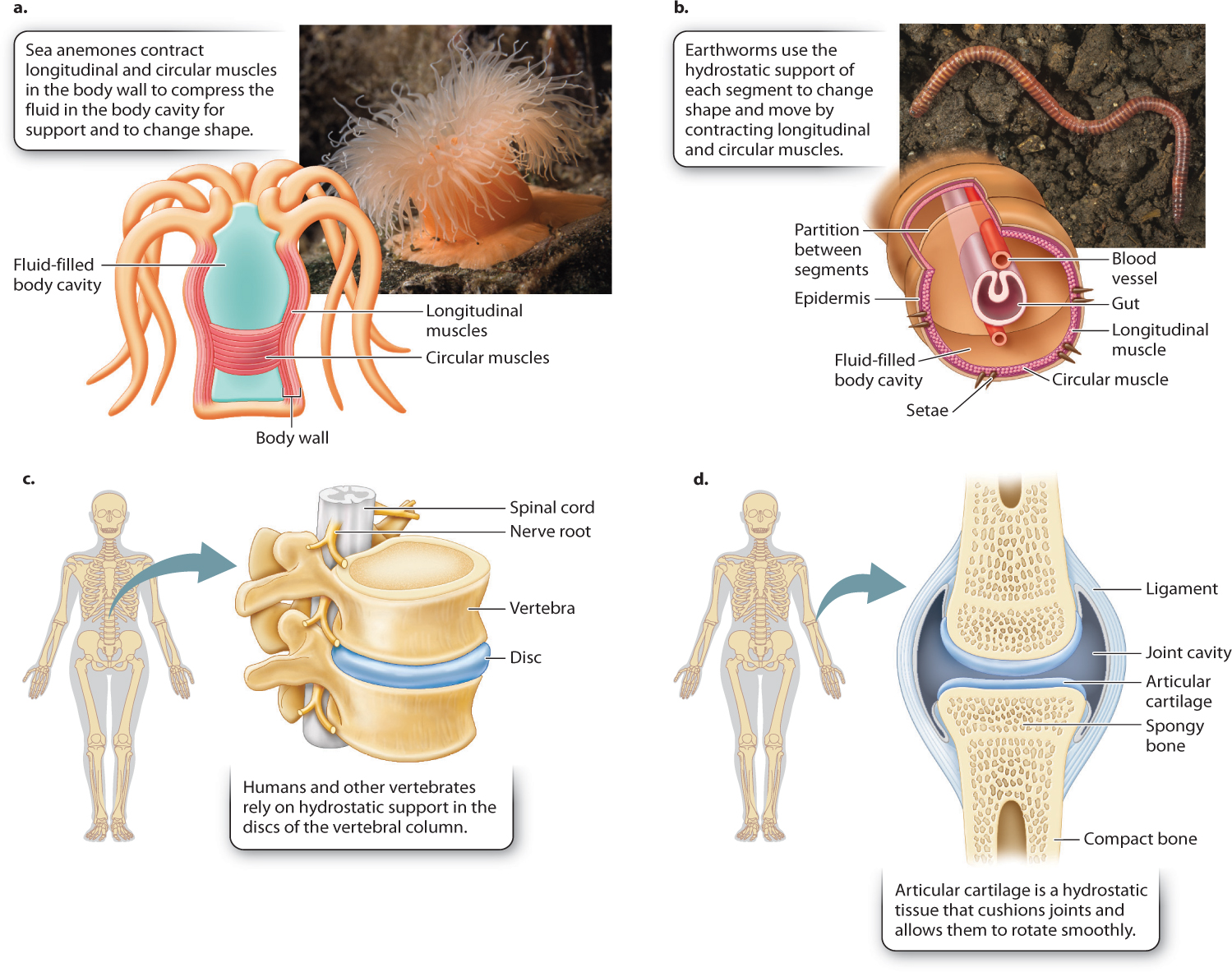
Similarly, earthworms burrow through the soil by moving a series of hydrostatic segments along their body (Fig. 37.15b). By contracting longitudinal muscles in a few segments, they shorten those segments and also widen them to anchor those segments against the soil. They then extend intervening segments between anchor points by contracting circular muscles, causing the segments to lengthen and move the animal’s body through the soil.
Because of their simple but effective organization, hydrostatic skeletons are well adapted for many uses. In groups such as squid and jellyfish, the hydrostatic skeleton has become specialized for jet-propelled locomotion. Large circular muscles contract to eject fluid from the body cavity, propelling the animal in the opposite direction. The muscles of other mollusks also employ a hydrostatic skeleton, allowing a clam, for example, to burrow into the sediment with a muscular “foot” and an octopus to adeptly manipulate its flexible tentacles. (Clams also have shells that form an exoskeleton to protect the body.) These same principles allow an elephant to control its trunk, and our tongue to manipulate food and assist in speech.
Even vertebrate animals with rigid endoskeletons have hydrostatic elements for flexibility and cushioning of loads transmitted by the skeleton. These include intervertebral discs and cartilage. Intervertebral discs are sandwiched between the bony vertebrae of the backbone (Fig. 37.15c). Each disc has a wall reinforced with connective tissue that surrounds a jelly-like fluid. Intervertebral discs enable the backbone to twist and bend. Disc walls damaged by repeated stress can rupture, causing significant lower back pain. Articular cartilage, another fluid-filled tissue, forms the joint surfaces between adjacent bones (Fig. 37.15d). The fluid within the cartilage resists the pressure between the ends of articulating bones, cushioning forces transmitted across the joint.
37.3.2 Exoskeletons provide hard external support and protection.
An exoskeleton is a rigid skeleton that lies external to the animal’s soft tissues. The first mineralized skeletons arose with sponges about 650 million years ago, but more successful exoskeletons subsequently evolved in other invertebrate groups adapted for life in aquatic and terrestrial environments (Fig. 37.16). Arthropod exoskeletons, which first evolved in aquatic crustaceans, protect animals from desiccation and physical insults. Consequently, exoskeletons were key to the success and diversification of insects in terrestrial environments about 450 million years ago. Because they are on the exterior of the animal, exoskeletons provide hard external support and protection, allowing muscles to attach from the inside. However, a main disadvantage is that exoskeletons limit growth.

Different invertebrates have different kinds of exoskeleton. The shells of bivalve mollusks such as clams and mussels form a hard, mineralized calcium carbonate exoskeleton (Fig. 37.16a and b). The calcium carbonate is reinforced by proteins and is therefore an example of a composite material, one that combines substances with different properties. Because of its composite nature, the shell is less brittle and more difficult to break than if it were made of just one substance. The shell expands as the organism grows, as epidermal cells deposit new layers of mineralized protein in regular geometric patterns. You can see the growth rings if you examine a mollusk shell. Although some marine bivalves achieve large size over many years, growth is limited by the rate at which new skeletal material can be deposited on the outside of the shell.
Arthropods, such as insects, have more complex exoskeletons formed of a cuticle that covers their entire body (Fig. 37.16c). The cuticle of insects is composed mainly of chitin, a nitrogen-containing polysaccharide. Marine crustaceans, such as crabs and lobsters, also incorporate calcium carbonate in their cuticle. The calcium carbonate makes the exoskeleton hard and stiff, helping to protect them from predators, much like the hardened cuticle of terrestrial insects. When initially formed, the cuticle of arthropods is soft, so the animal can grow before the cuticle is hardened. The cuticle remains flexible at joints, allowing motion between body segments. Mature cuticle consists of two layers. A thin outer, waxy layer minimizes water loss. Water loss is especially dangerous for arthropods, given their small size. The much thicker inner layer, whether flexible or stiff, is tough, making it hard to break.
Because a rigid exoskeleton restricts growth, arthropods shed their cuticle at intervals, a process termed molting. Molting allows arthropods to expand and grow before forming a new rigid exoskeleton. While offering several protective benefits, exoskeletons pose risks as well. Animals are vulnerable when the newly formed exoskeleton has not yet hardened. Exoskeletons are also hard to repair. If a skeleton is damaged while growing, the animal must produce an entirely new one. Because the exoskeleton is a thin-walled structure, it is prone to breaking if its surface area is very large. Consequently, the imperviousness of the monstrous insects depicted in science-fiction films is improbable—they would easily break from a blow to their exoskeleton.
37.3.3 The rigid bones of vertebrate endoskeletons are jointed for motion and can be repaired if damaged.
Endoskeletons first evolved in vertebrate animals about 500 million years ago, during the Cambrian explosion. This type of skeleton lies internal to most of an animal’s soft tissues. In contrast to exoskeletons, the bony endoskeletons of vertebrate animals can grow extensively and, when broken, can be repaired. Endoskeletons also provide protection for internal organs, such as the brain, lungs, and heart.
The bones of vertebrate skeletons consist of a variety of tubular, rodlike, and platelike elements that form a scaffold to which the muscles attach. Because bones are rigid and hard, there are joints between adjacent bones to enable movement. Muscles attach to the skeleton by connective tissue and specialized tendons made of collagen. Tendons transmit muscle forces, allowing the forces to be redirected and transmitted over a wide range of joint motion. Tendons, like a spring, also store and recover elastic energy.
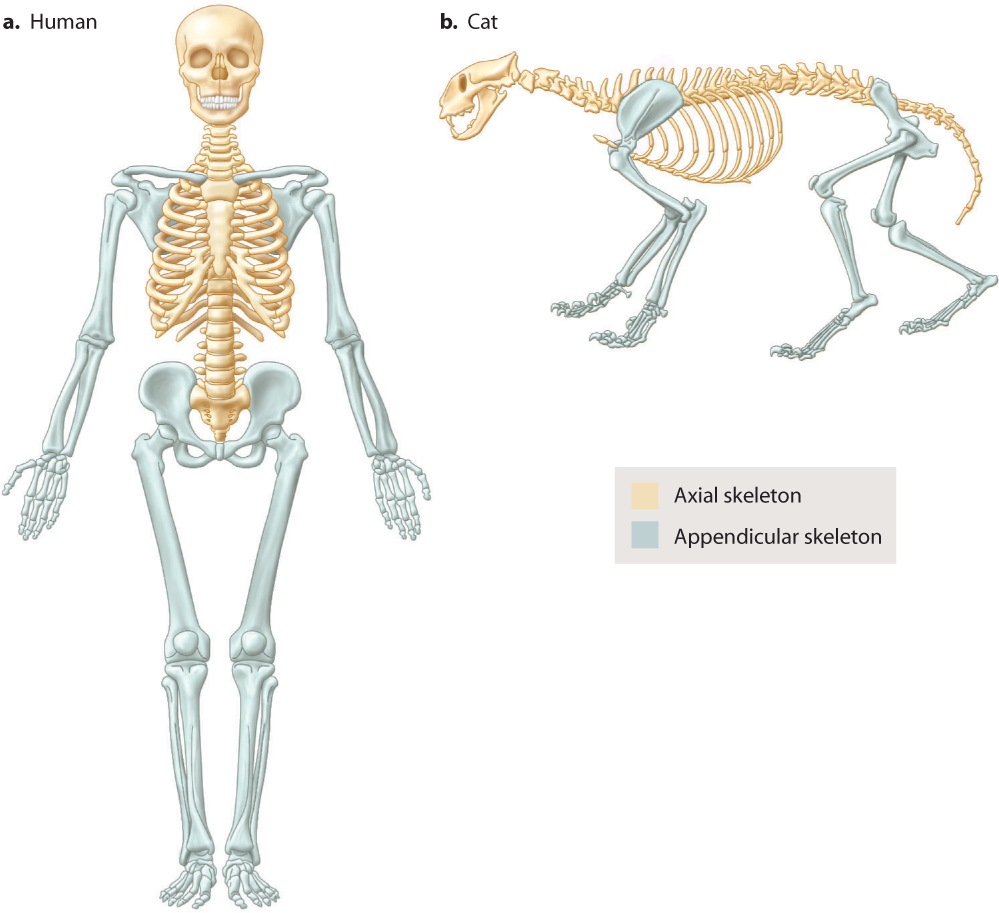
The vertebrate skeleton can be separated into axial and appendicular regions (Fig. 37.17). The axial skeleton consists of the skull and jaws of the head, the vertebrae of the spinal column, and the ribs. Bones of the skull protect the brain and sense organs of the head. The appendicular skeleton consists of the bones of the limbs, including the shoulder and pelvis. The axial and appendicular regions reflect the evolutionary ancestry of vertebrates. The axial skeleton formed first, to protect the head and provide support and movement of the animal’s body by bending its body axis. As a result, the axial skeleton is the main skeletal component of fishes. The appendicular skeleton comprises the pectoral and pelvic fins of fishes, which were elaborated into limbs when vertebrate animals first evolved to live on land. In a special group of lobed-fin fishes, the fin bones became adapted for an amphibious lifestyle, evolving into the limb bones that terrestrial vertebrates use for support against gravity and for movement on land.
Skeletal tissues have relatively few cells for their volume, similar to invertebrate exoskeletons. These tissues consist mostly of an extensive extracellular matrix that is secreted by specialized cells, forming a connective tissue external to the cells (Chapter 10). Three main extracellular tissues are produced in the formation of the vertebrate skeleton: bone, tooth enamel, and cartilage.
Bone tissue–forming cells called osteoblasts synthesize and secrete calcium phosphate as hydroxyapatite mineral crystals in close association with the protein collagen. Generally, bone consists of two-thirds hydroxyapatite mineral and one-third type I collagen protein. The combination of mineral and protein makes bone a composite material, like exoskeletons. Bone is rigid (because of the properties of the mineral), but also hard to break because it absorbs much energy before fracture (because of the properties of the collagen). The genetic bone disorder osteogenesis imperfecta leads to bones that are brittle and fragile because the osteoblasts produce insufficient collagen in the extracellular matrix. Bone gradually becomes more mineralized and brittle as a normal process of aging. Sharks and other elasmobranchs (the cartilaginous fishes) evolved unusual skeletons of calcified cartilage rather than mineralized bone tissue to achieve rigid mechanical support.
Articular cartilage located at the joint surfaces of a bone forms a gel-like matrix that resists fluid being squeezed out when forces press on the cartilage during movement. The cartilage is reinforced by type II collagen fibers, allowing the cartilage to cushion and distribute loads as the joint moves. Articular cartilage consists of 70% water, 15% large molecules called proteoglycans, and the remaining 15% collagen. Cartilage may degenerate for a variety of reasons, including rheumatoid arthritis and repetitive physical injury, but in general it lasts over an individual’s lifetime.
Quick Check 4
Glass is strong and rigid but can be easily broken. In contrast, a bone is strong and rigid but hard to break. How can this be?