38.2 PROPERTIES OF HORMONES
Hormones play diverse roles in the body, enabling regulation and coordination of multiple bodily functions in response to environmental cues. Because hormones are released into the bloodstream, they have the potential to affect many organ systems. Before discussing other important hormonal regulatory pathways, we must first consider how hormones target specific tissues and how different hormones exert their effect on different target cell types. Receptors on target organs respond to blood-borne hormones by binding the hormone, allowing the hormone to influence the function of specific cell targets. Certain classes of hormone (amine and peptide hormones) bind to receptors on the cell membrane and influence cellular function by signaling pathways (Chapter 9). The other general class of hormone (steroid hormones) is lipid soluble, so these hormones can diffuse across the cell membrane. Steroid hormones bind to receptors in the cytoplasm or in the nucleus and regulate transcriptional control of protein synthesis of the target cell.
38.2.1 Three main classes of hormone are peptide, amine, and steroid hormones.
Hormones are grouped into three general classes: hydrophilic peptide hormones, hydrophilic amine hormones, and hydrophobic steroid hormones, all of which are illustrated in Fig. 38.7. Peptide hormones and amine hormones are both derived from amino acids. Peptide hormones are short chains of amino acids, whereas amine hormones are derived from a single aromatic amino acid, such as tyrosine. All steroid hormones are derived from cholesterol. Whereas peptide hormones (which are sometimes large enough to be considered proteins) can evolve through changes in their amino acid sequence, steroid hormones cannot. Evolutionary changes in steroid hormone function depend instead on changes in the receptors they bind and the cellular responses they trigger.
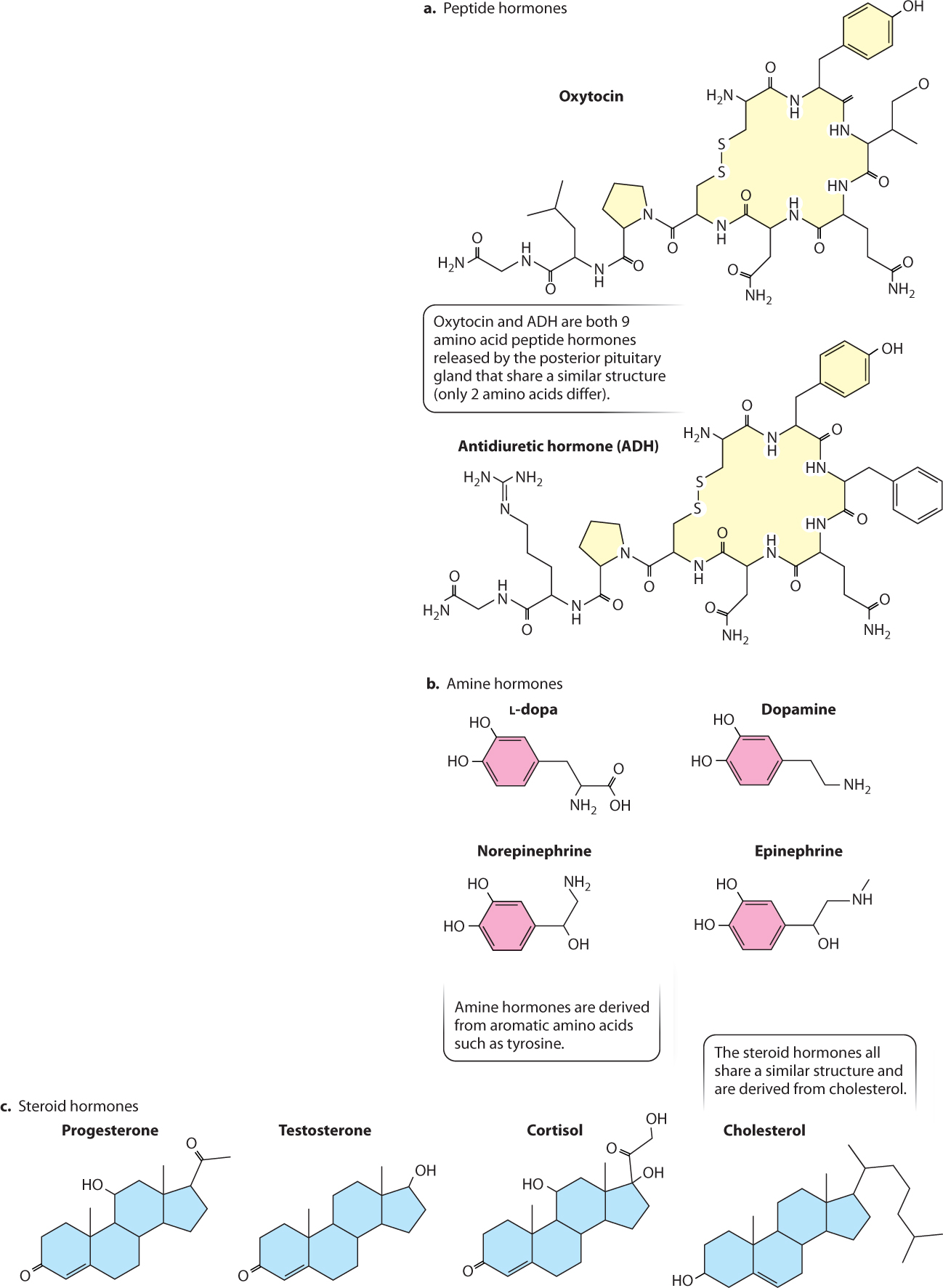
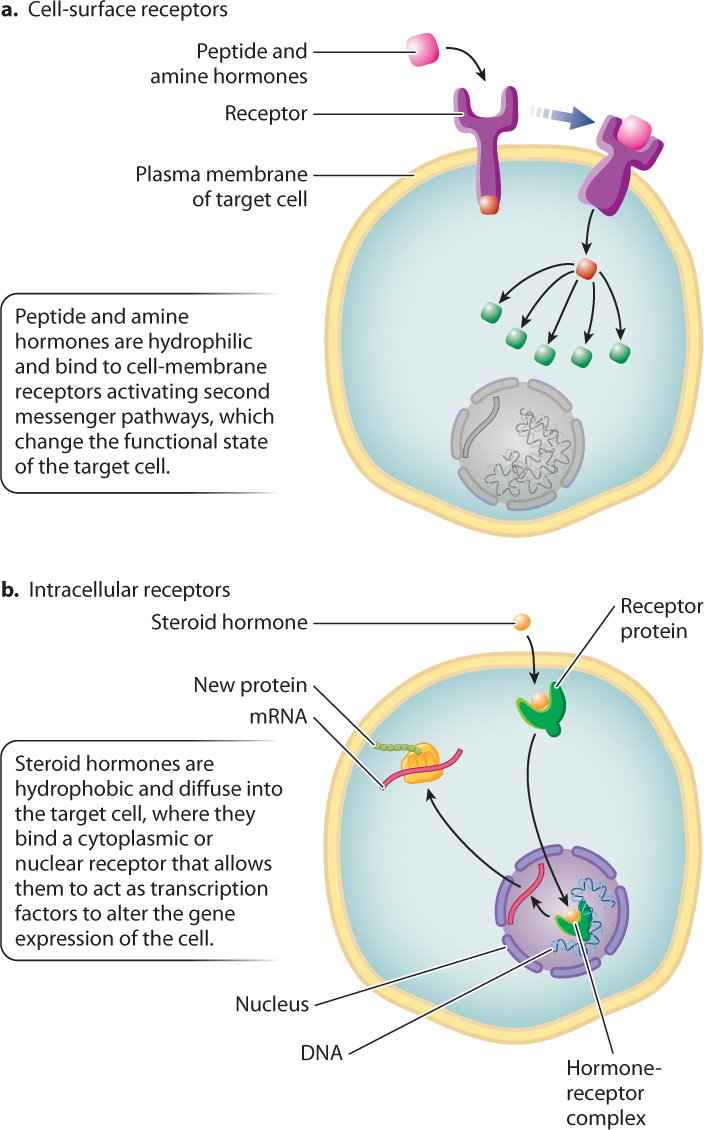
Peptide and amine hormones are more abundant than steroid hormones and are more diverse in their actions. Because most peptide and amine hormones are hydrophilic, they cannot diffuse across the plasma membrane. Instead, nearly all peptide and amine hormones bind to membrane receptors on the surface of the cell, triggering the intracellular second messenger pathways discussed in Chapter 9 (Fig. 38.8a). Peptide and amine hormones alter the biochemical activity of the target cell by activating or inactivating enzymes within the cell. Typically, they affect protein kinases (which phosphorylate other proteins), initiating signaling cascades within the target cell. These signaling cascades can lead to changes in gene expression or metabolism, trigger a cell to grow, divide, or change shape, or lead to the release of other hormones. Peptide and amine hormones act on timescales of minutes to hours.
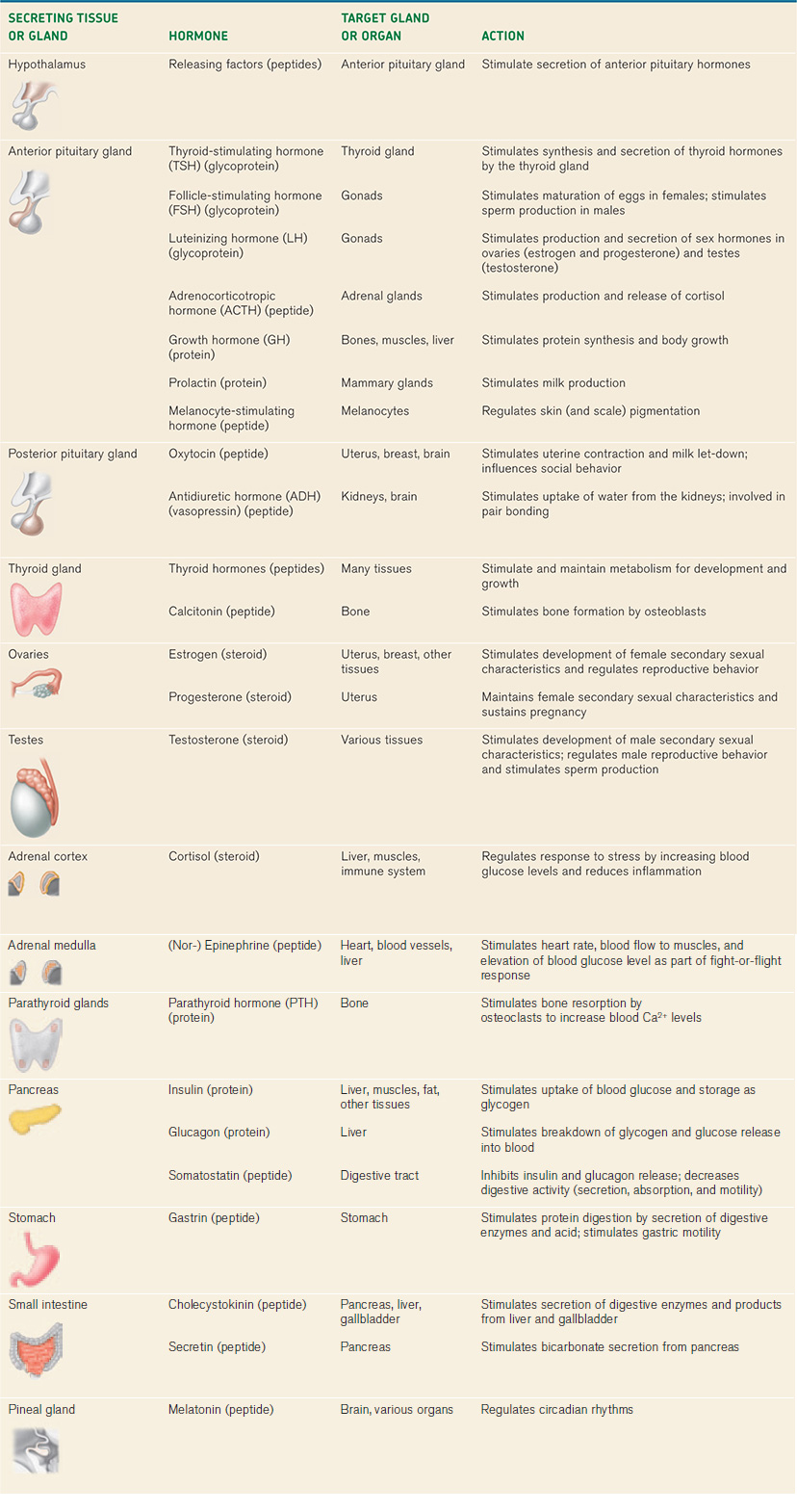
There are many examples of peptide hormones. In vertebrates, for instance, growth hormone stimulates protein synthesis and growth of many body tissues, particularly the musculoskeletal system (Table 38.2). As we saw, insulin and glucagon are peptide hormones released by the pancreas that regulate glucose metabolism by the liver, muscles, and other tissues. Gastrin and cholecystokinin are peptide hormones that regulate mammalian digestive function (Chapter 40). In addition to serving as a neurotransmitter released by the sympathetic nervous system (Chapter 35), epinephrine and norepinephrine (also known as adrenaline and noradrenaline) act as amine hormones, supporting an animal’s fight-or-flight response to stress.
In contrast to peptide and amine hormones, steroid hormones are hydrophobic. Therefore, they diffuse freely across the cell membrane to bind with receptors in the cytoplasm or nucleus (Fig. 38.8b), forming a steroid hormone–receptor complex. Hormone–receptor complexes that form in the cytoplasm are transported into the nucleus of the cell. These complexes most commonly act as transcription factors, stimulating or repressing gene expression and, therefore, altering the proteins produced by the target cell. Consequently, steroid hormones typically exert profound and long-lasting effects on the cells and tissues they target, with timescales of days to months. The actions of steroid hormones, therefore, tend to have a broader impact on body functions than those exerted by amine and peptide hormones.
The sex hormones estrogen, progesterone, and testosterone (Table 38.2), which we discuss in Chapter 42, are steroid hormones that regulate the differentiation, maturation, and functional state of the vertebrate reproductive organs. Cortisol is a steroid hormone that mediates a vertebrate animal’s response to stress and inhibits inflammation. Steroid hormones similar to cortisol are used medically to reduce the symptoms of certain inflammation disorders. However, they carry risks because they also depress the immune system, increasing the chance of infection and diminishing wound healing.
38.2.2 Hormonal signals are typically amplified.
Hormones are typically released in small amounts. So how is it possible for them to have large effects on the overall physiology of an organism? The answer is that hormone signals are amplified. Amplification occurs through a series of signaling steps between different endocrine glands in a hormonal pathway, as well as signal transduction steps within the target cells after the hormone binds to a cell receptor. Signals are thus amplified at each step of the pathway, resulting in a large effect on the target organ system.
Hormonal signaling pathways between endocrine glands and tissues are often referred to as endocrine axes. For example, in vertebrates, the hypothalamic–pituitary axis involves hormonal signals that are amplified along a pathway from the hypothalamus to the anterior pituitary gland, and then from the anterior pituitary gland to target glands or tissues in the body.
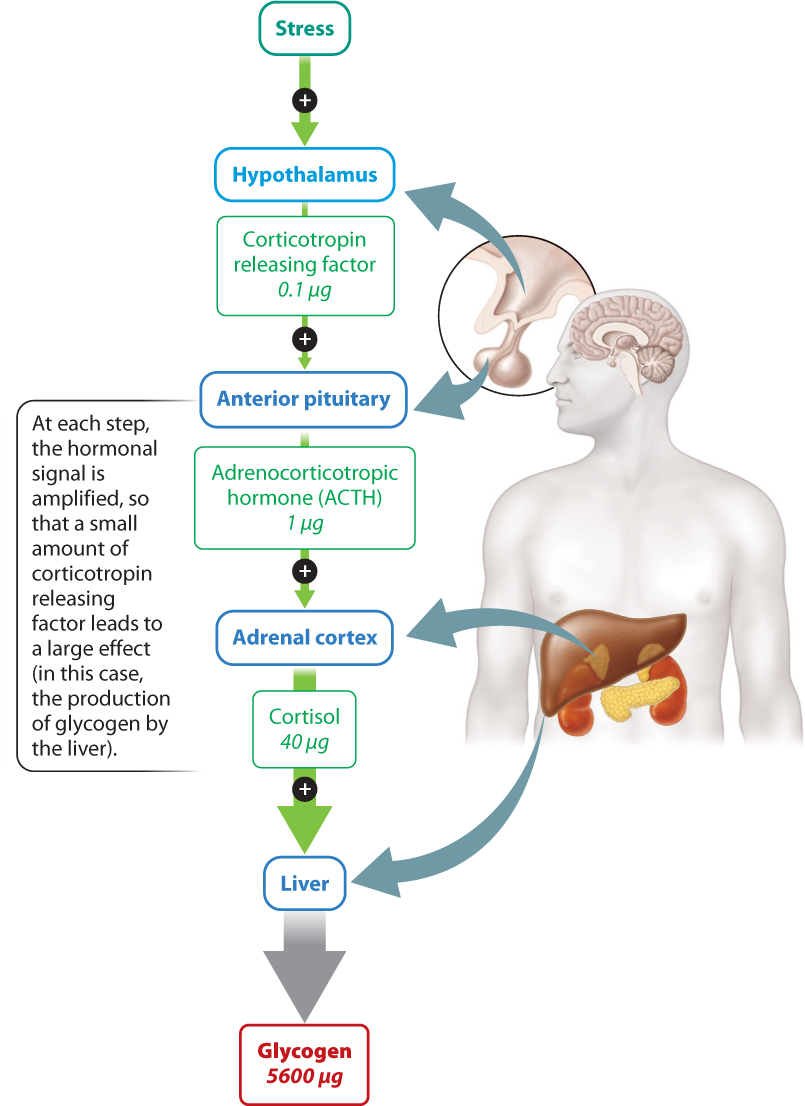
The vertebrate hypothalamic–pituitary axis begins with trace amounts of peptide hormones called releasing factors that are initially released by the hypothalamus (Fig. 38.9). They signal to the anterior pituitary gland, leading to a much larger release of associated hormones from that organ. For example, corticotropin releasing factor secreted by the hypothalamus stimulates the release of a larger amount of adrenocorticotropic hormone (ACTH) by the anterior pituitary gland.
Hormones released by the anterior pituitary gland in turn bind cell receptors in the target organ. In this case, ACTH acts on cells of the adrenal cortex, stimulating their secretion of the hormone cortisol. Cortisol acts on many different cells and tissues in the body, causing what is known as an acute stress response. Among its effects, it causes the liver to convert glucose to glycogen (although through effects on other cells it also increases blood glucose levels). This action yields 56,000x more glycogen molecules than the initial number of releasing factors secreted by the hypothalamus.
Similar signaling amplification cascades occur in invertebrates in the regulation of insect molting and metamorphosis (see Fig. 38.3). The amount of brain hormone that is produced is dwarfed by the amount of ecdysone or juvenile hormone that is released to regulate the growth and metamorphosis of the insect’s body. As in vertebrates, second messenger signaling cascades within receptor cells of the body respond to the release of these hormones by amplifying the hormone signal in the cell. Thus, amplification by signal transduction pathways applies both to second messenger systems within a cell as well as the chemical signals that are transmitted as hormones between glands and tissues in an organism’s endocrine system.
Quick Check 2
Why do steroid and peptide hormones bind different kinds of receptor and how does this difference affect the resulting signaling pathways in the target cell?
38.2.3 Hormones act specifically on cells with receptors that bind the hormone.
Hormones are chemical signals that enable communication between different cells in an organism. Because hormones are secreted into the bloodstream, they achieve specificity by binding to receptors on the surface of or inside the target cells. Therefore, it is the presence or absence of a receptor for a given hormone that determines which cells respond and which ones do not. For example, when the hormone oxytocin is released into the bloodstream of mammals, it affects only cells that express a receptor on the cell surface capable of binding oxytocin as it flows by in the bloodstream. These cells are uterine muscle cells and secretory cells in breast tissue. When oxytocin binds these cell-surface receptors, uterine muscle cells are stimulated to contract, and secretory cells in breast tissue release milk during breastfeeding. The effect of oxytocin also depends on cells of other organs not expressing the receptor so that the hormone can exert its effect on these specific tissues.
The binding of a hormone to its receptor leads to changes in the target cell, resulting in a cellular response. The specific action of a hormone depends on the kind of response that it triggers in the target cell. For example, the binding of a hormone can lead to changes in ion fluxes across the cell membrane, activation of an intracellular signal transduction cascade, or changes in gene and protein expression.
Quick Check 3
What general features make a chemical compound a hormone, and how do hormones achieve specificity for certain kinds of target cells?
38.2.4 Hormones are evolutionarily conserved molecules with diverse functions.
Most hormones have an ancient evolutionary history. The structures of many hormones are evolutionarily conserved. Some vertebrate hormones can also be found in many invertebrates, but typically they serve different functions. Since the first vertebrate animals diverged from invertebrates over 500 million years ago, this indicates that some vertebrate hormones are even older. And, in many cases, their roles in many invertebrate animals have yet to be discovered.
Thyroid-stimulating hormone (TSH), a hormone released by the anterior pituitary gland that targets the thyroid gland, regulates metabolism in vertebrate animals but triggers metamorphosis in amphibians and feather molt in birds. It has even been found in snails and other invertebrates that lack a thyroid gland. Its function in snails appears to be to stimulate the number of sperm or eggs produced.
Recent genomic analysis has also revealed that the receptors for many hormones evolved well before the hormones with which they now interact. Even though the structure of a hormone or its receptor is often largely unchanged across diverse groups of organisms, hormones and their receptors can readily be selected to take on new roles. An example is the shift in function of TSH to regulate metabolism in mammals and feather molt in birds from an earlier role stimulating sperm or egg production in snails. Thus, the biochemical evolution of hormones and their target cell receptors can be readily altered as organisms evolve new behaviors and exploit new environments.
Another intriguing finding is that many peptides originally identified as hormones in various tissues have also been found to function as neurotransmitters in the nervous system. For example, oxytocin, which stimulates uterine contractions and the release of milk, also serves as a neurotransmitter in the brain and is believed to influence social behavior, as well as stimulate sexual arousal in mammals. Similarly, another peptide hormone, antidiuretic hormone, which regulates water uptake in the kidneys (Chapter 41), also functions as a neurotransmitter in the brain influencing mammalian mating and pair-bonding behavior. Hence, the same compound, expressed in two different cell types, may have entirely different functions. The roles of hormones as chemical messengers are varied within an organism and easily changed over the course of evolution.