39.4 CIRCULATORY SYSTEMS
The O2 carried by hemoglobin in red blood cells is transported to tissues throughout the body by the circulatory system. The closed circulatory systems of animals are made up of a set of internal vessels and a pump—the heart—to transport the blood to different regions of the body. Closed circulatory systems have two conflicting requirements: They must produce enough pressure to carry the circulating blood to all the tissues, yet once the blood reaches smaller vessels that supply the cells within the tissues the blood pressure and flow rate must not be too high. High pressure would push fluid through the walls of the smaller vessels and cause the blood to flow so quickly that there would not be enough time to exchange gases.
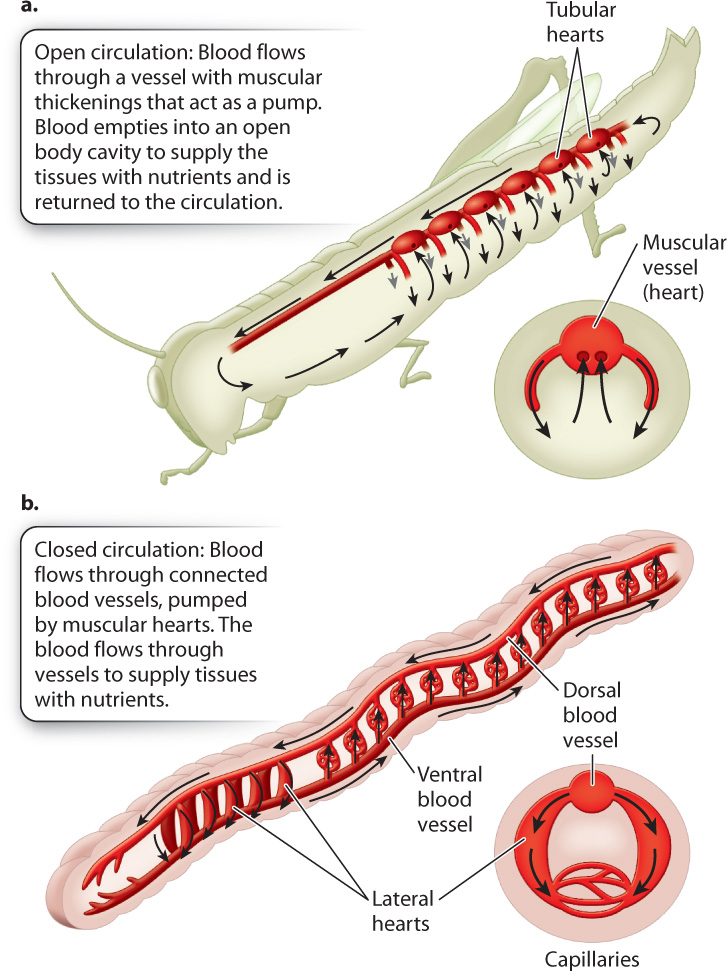
Generally, smaller animals, such as insects and many mollusks, have open circulatory systems that contain few blood vessels: Most of the circulating fluid, the hemolymph, is contained within the animal’s body cavity. The hemolymph bathes the animal’s tissues and organs (Fig. 39.15a). Open circulatory systems have limited control of where the fluid moves. Muscles that are active in locomotion can assist circulation of the hemolymph, and some invertebrates have simple hearts with openings that help pump fluid between different regions of the animal’s body cavity.
Animals with closed circulatory systems tend to be larger, although segmented worms have evolved closed circulatory systems that allow them to expand individual body segments to burrow underground (Fig. 39.15b). The first cephalopods (squid and octopus), which have closed circulatory systems, evolved about 480 million years ago and were early successful predators in Paleozoic oceans. A closed circulatory system delivers O2 at high rates to exercising tissues, providing them with the energy used to chase prey. In contrast, open circulatory systems generally operate under low pressure and have limited transport capacity. As a result, animals with open circulatory systems are less active. Their ability to control the delivery of respiratory gases and metabolites to specific tissues and regions is also limited. However, insects can be very active despite having an open circulatory system because they obtain O2 by their tracheal system independently of the circulatory system.
Closed circulatory systems can control blood flow to specific regions of the body by varying the resistance to flow. For example, wading birds reduce blood flow to their legs when they are in cold water to reduce heat loss, and vertebrates can increase blood flow to their muscles to deliver O2 and nutrients during exercise. In order to pump blood through a set of closed interconnected vessels, a muscular heart is needed to produce sufficient pressure to overcome the flow resistance of the vessels.
39.4.1 Circulatory systems have vessels of different sizes to facilitate bulk flow and diffusion.
For a fluid like blood to flow through a set of pipes, pressure (P) is required to overcome the resistance (R) to flow. The pressure developed by a heart when it contracts establishes a rate of blood flow that is governed by P/R. That is, the rate of blood flow increases with an increase in pressure and decreases with an increase in resistance. The resistance to flow is determined in part by the fluid’s stickiness (its viscosity) and the vessel’s length. Longer vessels of a given size impose greater resistance. However, the main factor governing resistance to flow is the vessel’s radius (r). Resistance is proportional to 1/r4, which means that if a vessel’s radius is reduced by half, its resistance to flow increases 16 times.
The high resistance of narrow vessels presents a challenge. To keep diffusion distances short, gas exchange between cells and blood requires narrow vessels. How does the circulatory system overcome the dramatic increase in resistance with narrower diameters? Animal circulatory systems are organized to have blood flow over longer distances in a relatively few large-diameter vessels with low resistance to flow (Fig. 39.16a).
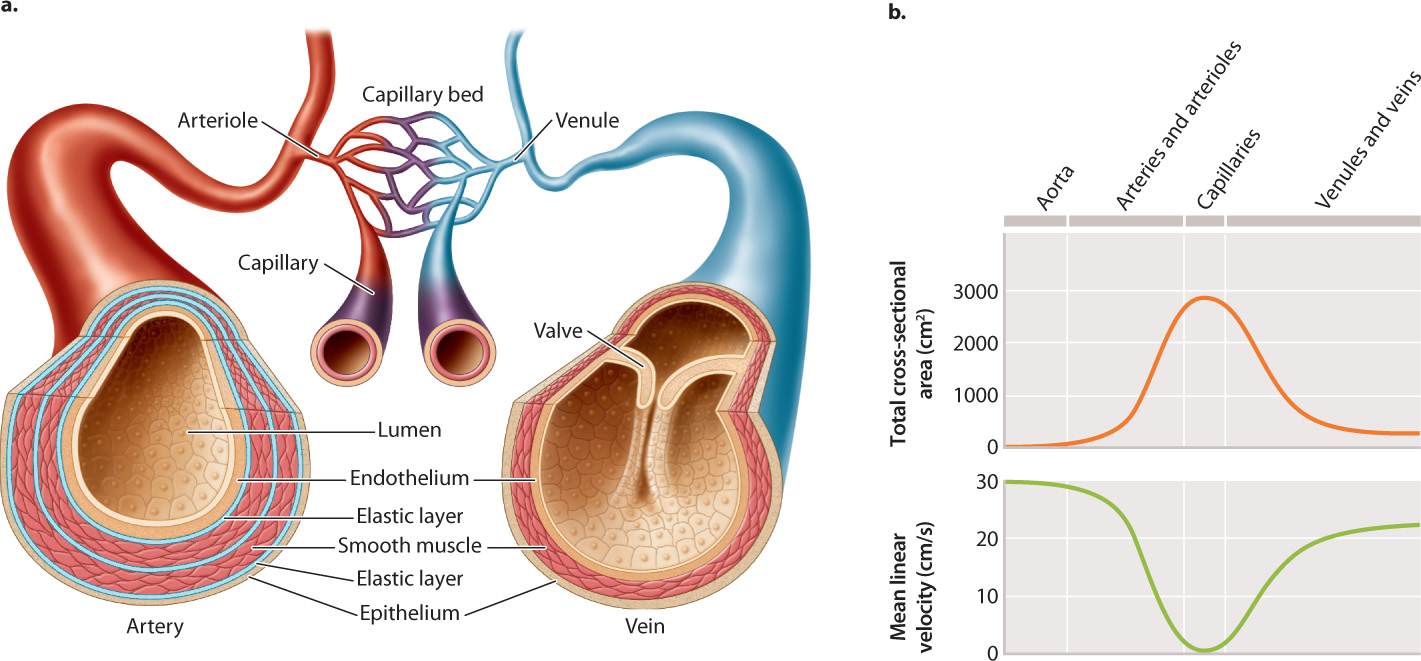
Arteries are the large, high-pressure vessels that move blood flow away from the heart to the tissues. Veins are the large, low-pressure vessels that return blood to the heart. Arteries branch into blood vessels of progressively smaller diameter called arterioles, and these arterioles ultimately connect to finely branched networks of very small blood vessels called capillaries. It is at the capillaries that gases are exchanged by diffusion with the surrounding tissues. The number of smaller-diameter vessels at each branching point in the circulatory system greatly exceeds the number of larger diameter vessels. The reverse organization is found on the return side of circulation: Numerous capillaries drain into vessels of progressively larger diameter called venules, and the venules drain into a few larger veins that return blood to the heart.
This organization has two advantages. First, it maintains the same volume of blood flow at all levels within the circulatory system: The increased resistance to flow in the capillaries is offset by the large increase in the number of capillaries. Second, it enables the blood to flow more slowly in smaller vessels (Fig. 39.16b), providing time for gases and metabolites to diffuse into and out of the neighboring cells. Indeed, the diameter of a capillary is so small that red blood cells pass through one at a time, and must be flexible to do even that.
39.4.2 Arteries are muscular, elastic vessels that carry blood away from the heart under high pressure.
When an animal becomes active after a period of rest and feeding, blood flow must be increased to its muscles and reduced to its digestive organs (Fig. 39.17). How do animals accomplish this? The answer comes from physics: the strong effect of vessel radius on flow resistance. Arterioles supplying regions of the body that need less blood shrink in diameter by contracting the circular smooth muscle fibers in their wall. The reduction in diameter increases the resistance to flow and thereby reduces the rate of blood flow. For example, when you put your hands or feet into cold water, the arterioles contract, reducing the loss of heat from blood flowing to your hands and feet.
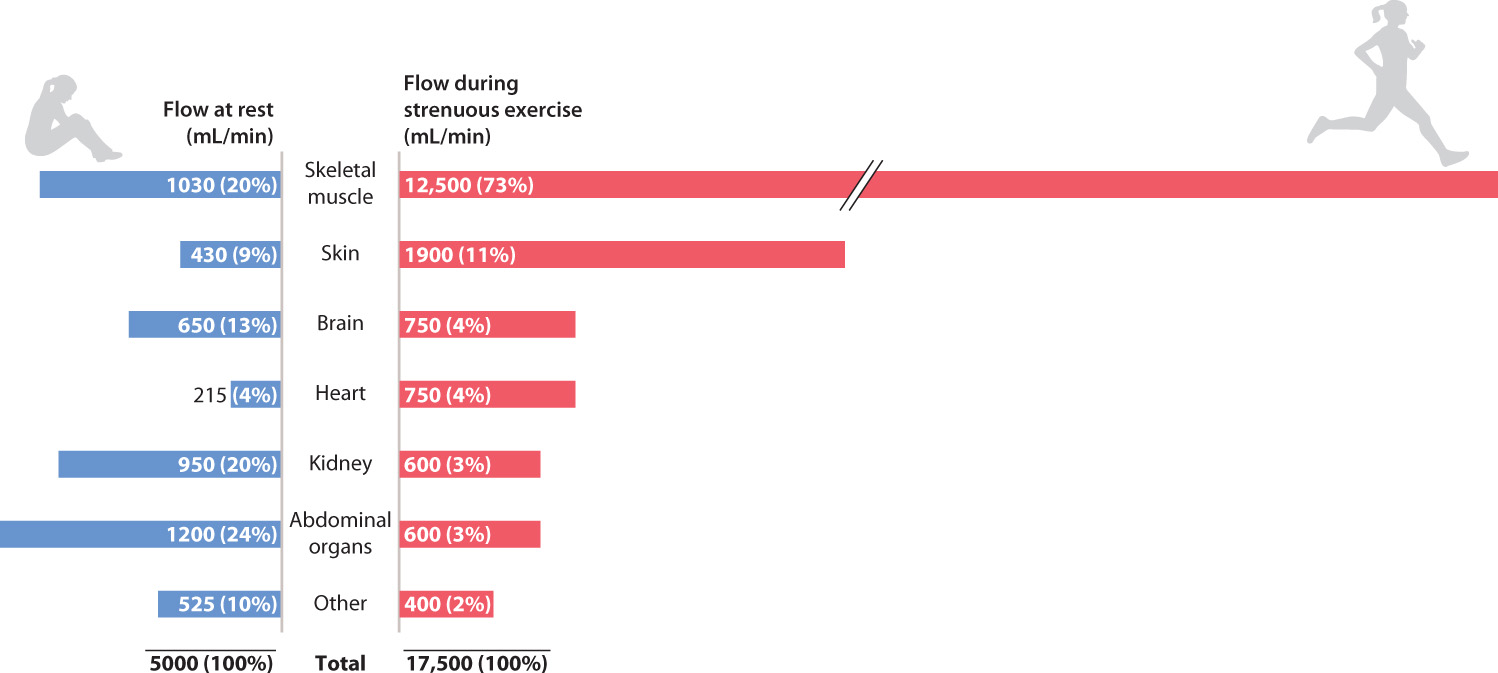
Arterioles supplying regions of the body that need more blood expand in diameter by relaxing their smooth muscles. The larger vessels offer less resistance, so more blood can flow through them. What is the mechanism that causes the smooth muscles to relax when cells in neighboring tissue become more active? As the rate of cellular respiration in these cells increases, they begin releasing more CO2 as well as other metabolites, such as lactate and hydrogen ions, that are produced in respiration. The increase of these molecules in the bloodstream acts as a signal to the smooth muscle to begin relaxing. In this way, blood flow increases to the capillaries to match O2 and nutrient delivery to the metabolic need of the cells.
A strong pulse of flow pushes against the arterial walls about every second in a human as the heart beats to pump blood through the arteries. This pulse of flow causes a momentary expansion of the arteries. Artery walls are able to withstand these repeated pressure pulses because they contain multiple elastic layers composed of two proteins: collagen and elastin. Collagen fibers are strong and resist the expansion of the arterial wall during these pressure pulses. Together with elastin fibers, the collagen provides an elastic rebound of the arterial wall once the pulse has passed, returning energy to help smooth out blood flow. The collagen and elastin resist overexpansion of the arterial wall at local sites. If an artery wall deteriorates, the collagen and elastin can become so thin that the artery bulges outward. The outward bulge, called an aneurysm, can lead to a life-threatening rupture.
Quick Check 5
What change in vessel shape most affects resistance to blood flow?
39.4.3 Veins are thin-walled vessels that return blood to the heart under low pressure.
Blood collected from local capillary networks returns to the heart through progressively larger veins that ultimately drain into the two largest veins, the venae cavae (singular, vena cava). The venae cavae drain blood from the head and body into the heart. There is little pressure available to push the blood forward in the veins because pressure has been lost to the resistance of the arterioles and capillaries. Consequently, veins are thin walled and have little smooth muscle or elastic connective tissue. Because of the low pressure, blood tends to accumulate within the veins: As much as 80% of your total blood volume resides in the venous side of your circulation at any one time. When you stand or sit for a long time, blood may pool within the veins of your limbs.
Veins located in the limbs and in the body below the heart have one-way valves that help prevent blood from pooling due to gravity (see Fig. 39.16a). However, the most important aid to returning blood to the heart is the voluntary muscle contraction that occurs during walking and exercise, which exerts pressure on the veins. Above the level of the heart, the return of blood to the heart through the veins is assisted by gravity.
39.4.4 Compounds and fluid move across capillary walls by diffusion, filtration, and osmosis.
We have seen that O2 and CO2 move between capillaries and tissues by diffusion. In addition, water, certain ions, and other small molecules, but not proteins, move from capillaries into the surrounding interstitial fluid (the extracellular fluid surrounding vessels) by filtration, forced by blood pressure.
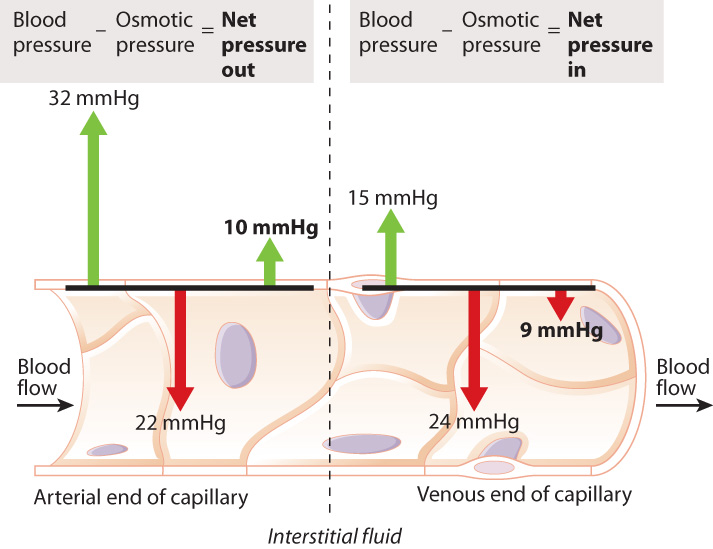
Why doesn’t the blood plasma lose all its water and ions over time? As water is filtered out of the capillaries, some ions, proteins, and other large compounds remain inside, increasing in concentration (Fig. 39.18). As a result, there is a tendency of water to flow back into the capillary by osmosis (Chapter 5). At the same time, blood pressure decreases because of the flow resistance imposed by the capillaries. When the pressure is no longer high enough to counter osmosis, water moves back into the capillaries.
In vertebrates, excess interstitial fluid is also returned to the bloodstream by means of the lymphatic system, a network of vessels distributed throughout the body. The fluid that enters the lymphatic system is called the lymph. Small lymphatic vessels merge with progressively larger thin-walled vessels, draining the lymph into lymphatic ducts, which empty into the venous system and then the heart. Lymphatic vessels have one-way valves, similar to those in the veins, that assist the return of lymph to the circulatory system. Local muscle contractions also help to pump the lymph toward the thoracic duct. In addition to its role in returning plasma to the bloodstream, the lymphatic system has important functions in the immune system (Chapter 43).
Case 7 Predator-Prey: A Game of Life and Death
39.4.5 How do hormones and nerves provide homeostatic regulation of blood flow as well as allow an animal to respond to stress?
An animal that is dehydrated or has lost blood following an injury may experience a prolonged drop in blood pressure. In such cases, the posterior pituitary gland releases antidiuretic hormone (ADH), also known as vasopressin (Chapter 38), into the circulation. ADH causes the arteries to contract, increasing their resistance to blood flow. The higher resistance elevates blood pressure throughout the body.
Even under less stressful conditions, blood pressure is continually being adjusted to remain within normal physiological limits, another example of homeostasis. The autonomic nervous system plays an important role in keeping blood pressure at healthy levels (Chapter 35). If blood pressure in the general circulation drops, animals reduce the supply of blood to the limbs by constricting arteries that supply the limbs. This response helps maintain blood pressure to the heart, brain, and kidneys. In this case, sympathetic neurons synapsing on the smooth muscles of arterioles in the limbs stimulate these muscles to contract. In contrast, when blood pressure is too high, sympathetic neurons that synapse on the smooth muscles are inhibited. The smooth muscles relax, reducing resistance in the arteries and increasing blood flow. These two responses are called vasoconstriction and vasodilation.
In some circumstances, it may be advantageous for an animal’s blood pressure to rise higher than normal levels. One example is during the fight-or-flight response that occurs when animals are faced with a stressful situation, as when prey animals are alerted to the presence of a predator, and when predators locate a potential prey. The sympathetic nervous system stimulates the adrenal gland to release the hormone adrenaline (also called epinephrine). The surge of adrenaline into the bloodstream— the “adrenaline rush”—causes an animal to become alert and aroused. At the same time, the sympathetic nervous system acts to increase heart rate and respiratory rate and inhibit digestive processes. Arteries supplying limb muscles dilate to enhance blood flow to exercising muscles, while arteries supplying organs within the digestive system constrict. The increase in heart and breathing rates increases the rate of O2 delivery to the exercising muscles to meet their increased demand for ATP. These physiological changes, along with a heightening of sensory acuity, prepare an animal for the more intensive activity needed to chase or flee.
In contrast, when an animal has recently eaten a meal, its sympathetic nervous system is inhibited and its parasympathetic nervous system is stimulated, increasing digestive secretions and the activity of gut smooth muscle (Chapter 40). Thus, local regulation of blood flow within different regions and organ systems of the body is integrated with more general feedback from the nervous and endocrine systems. The result is homeostatic regulation of blood pressure that monitors digestion and meets the need for O2 delivery to all metabolizing cells of the animal. These shifts in blood flow are readily controlled by vasoconstriction or dilation of the arteries supplying the different organs.