39.5 THE EVOLUTION, STRUCTURE, AND FUNCTION OF THE HEART
The pressure required to drive blood through blood vessels is generated by the heart. Animal hearts are pumps made of muscle that rhythmically contract, producing this pressure. Early hearts were as simple as a muscular thickening of a small region of a blood vessel. Such hearts are still found in insects (see Fig. 39.15a). More complex hearts have chambers that expand and fill with deoxygenated blood returning from the animal’s tissues. After filling, the heart muscle contracts, pumping deoxygenated blood to lungs or gills and newly oxygenated blood onward to the animal’s tissues. The closed circulatory system of cephalopod mollusks has three hearts—two that pump deoxygenated blood through the gills and another that pumps oxygenated blood to the body. In contrast, vertebrates have a single heart with at least two chambers, one for receiving blood and the other for pumping blood to the body during each heartbeat cycle. Hearts also have valves to ensure that blood does not flow backward when the heart contracts.
A theme in the evolution of vertebrate circulatory systems is the progressive separation of circulation to the gas exchange organ from the circulation to the rest of the body. In fish, deoxygenated blood is first pumped to the gills to gain O2 before flowing to the rest of the body and then returning to the heart. This single circulation path limits the rate of blood flow to metabolically active body tissues: The resistance imposed by the small vessels in the gills slows the blood flow considerably before it even reaches the tissues. By contrast, birds and mammals evolved a separate pulmonary circulation to the lungs and systemic circulation to the rest of the body. The evolution of separate pulmonary and systemic circulations was made possible by the evolution of a four-chambered heart. This organization has two advantages: It increases the supply of oxygenated blood to active tissues, and it increases the uptake of O2 at the gas exchange surface.
39.5.1 Fish have two-chambered hearts and a single circulatory system.
Fish hearts have two chambers (Fig. 39.19), the atrium and ventricle. Deoxygenated blood returning from the fish’s tissues enters the atrium, which fills and then contracts to move the blood into a thicker-walled ventricle. The muscular ventricle pumps the blood through a main artery to the gills for uptake of O2 and elimination of CO2. Oxygenated blood collected from the gills travels to the tissues through a large artery called the aorta.
The small gill capillaries impose a large resistance to flow. As a result, much of the blood pressure is lost in moving blood through the gills. This loss of pressure limits the flow of oxygenated blood to body tissues.

39.5.2 Amphibians and reptiles have three-chambered hearts and partially divided circulations.
As animals moved onto land, the transition from breathing water to breathing air had important consequences for the organization of vertebrate circulatory systems. Land vertebrates evolved hearts that separated the circulation of deoxygenated blood pumped to their gas exchange organs from circulation of oxygenated blood delivered to their body tissues. Gas exchange became more efficient and O2 delivery increased. Metabolic rates rose, and animals became capable of greater activity.
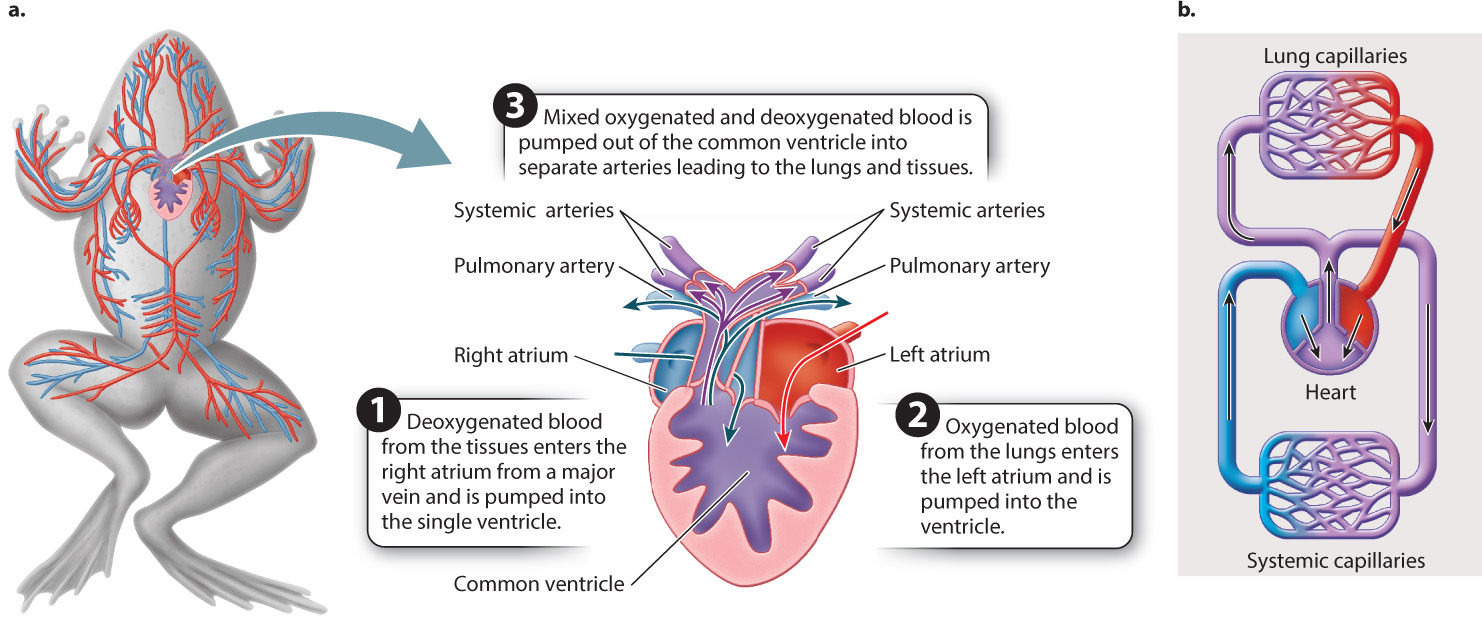
Reflecting their lifestyle, both aquatic and terrestrial, amphibians evolved a variety of ways to breathe. Some retain gills, while others use lungs or their skin, or some combination of these, to breathe. Amphibians evolved an additional atrium, dividing their heart into two separate atria and a single ventricle (Fig. 39.20). This arrangement partially separates the pulmonary and systemic circulations, so in amphibians freshly oxygenated blood can be pumped under higher pressure to the body than is the case in fishes. However, because amphibians have a single ventricle, oxygenated blood returning to the heart from the gills or lungs mixes with deoxygenated blood returning from the animal’s body before being pumped from the ventricle. Anatomical features of the ventricle help to limit this mixing, but nonetheless mixing still hinders the supply of oxygenated blood to an amphibian’s body.
Most reptiles also have a three-chambered heart, but their single ventricle has internal ridges to improve the separation of deoxygenated and oxygenated blood. As a result, reptiles are able to deliver more O2 to their tissues than amphibians can. At the same time, the incompletely divided ventricle provides reptiles with versatile circulatory responses during exercise and when diving underwater. For example, when they dive, the blood of turtles and sea snakes bypasses their lungs, which they cannot use underwater.
39.5.3 Mammals and birds have four-chambered hearts and fully divided pulmonary and systemic circulations.
Mammals and birds evolved four-chambered hearts with separate atria and separate ventricles (Fig. 39.21). This arrangement completely separates blood flow to the lungs from blood flow to the tissues. As a result, these animals can pump blood to their lungs under lower pressure, allowing increased uptake of O2, while at the same time supplying blood gases and nutrients to their tissues at high pressure. Whereas the systemic blood pressure of fish is only a few mmHg, in a bird or mammal it is 100 mmHg or higher. Moreover, blood flowing to the tissues is fully oxygenated at the outset, delivering more O2 to body tissues at a higher rate. This change in heart structure and circulatory pattern was linked to an increase in the metabolic rates and the evolution of endothermic lifestyles in birds and mammals (Chapter 40).
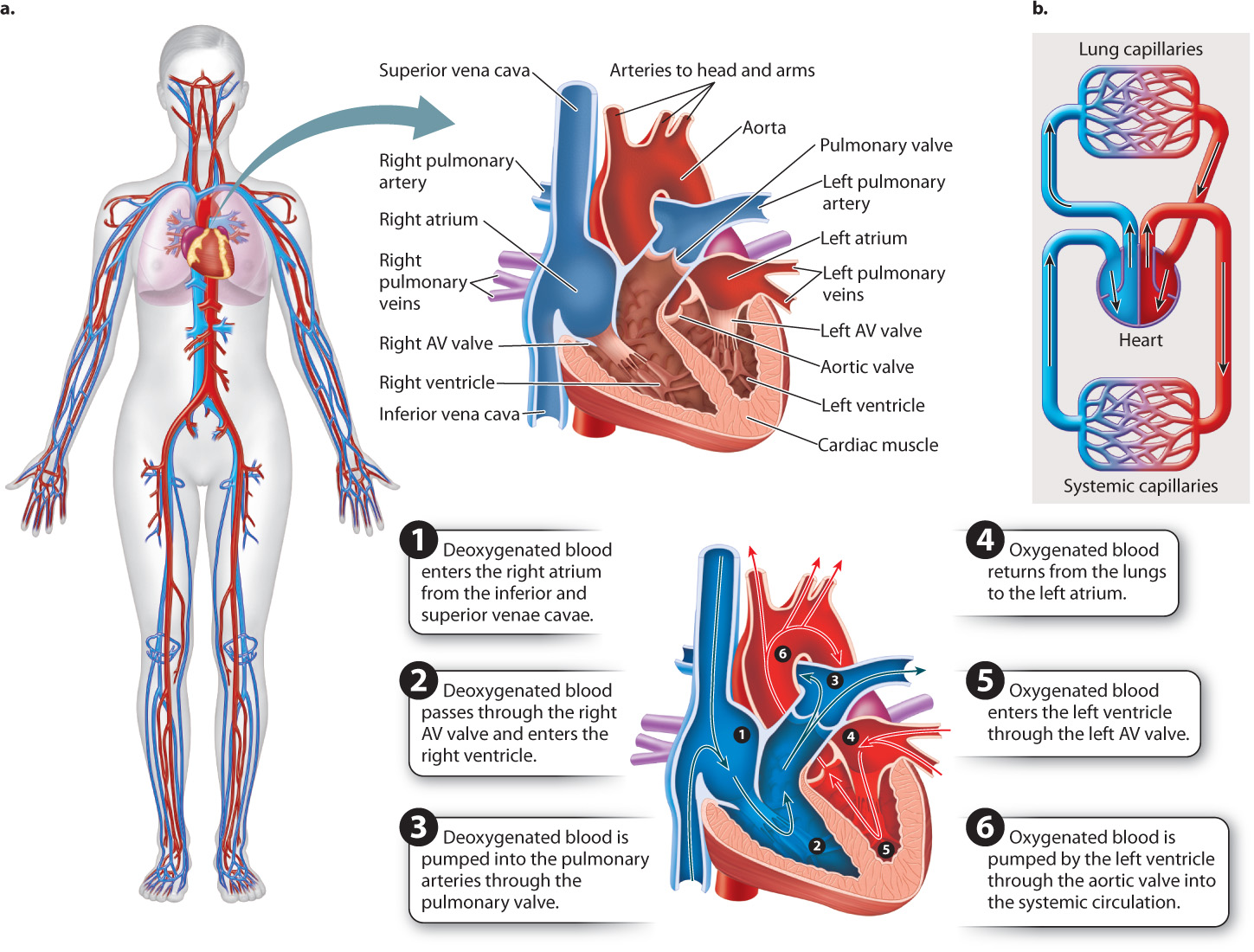
Following the pattern established in amphibians and reptiles, deoxygenated blood enters the right atrium from the venae cavae and, when the atrium contracts, moves through an atrioventricular (AV) valve into the right ventricle (Fig. 39.21). When the right ventricle contracts, blood is pumped through the pulmonary valve into the pulmonary trunk, which divides into the left and right pulmonary arteries, and then to the lungs for oxygenation. The walls of the right ventricle are thinner than those of the left ventricle, and its weaker contractions eject blood at a lower pressure. As a result, the blood of the pulmonary circulation moves at a slower rate, allowing greater time for gases to diffuse into and out of the lungs.
The oxygenated blood returns from the lungs through the pulmonary veins and enters the left atrium of the heart. When the left atrium contracts, blood is pumped through a second atrioventricular valve into the left ventricle. The thick muscular walls of the left ventricle eject the blood under high pressure to the body. Oxygenated blood leaving the left ventricle passes through the aortic valve and then flows to the head and the rest of the body through a large artery called the aorta.
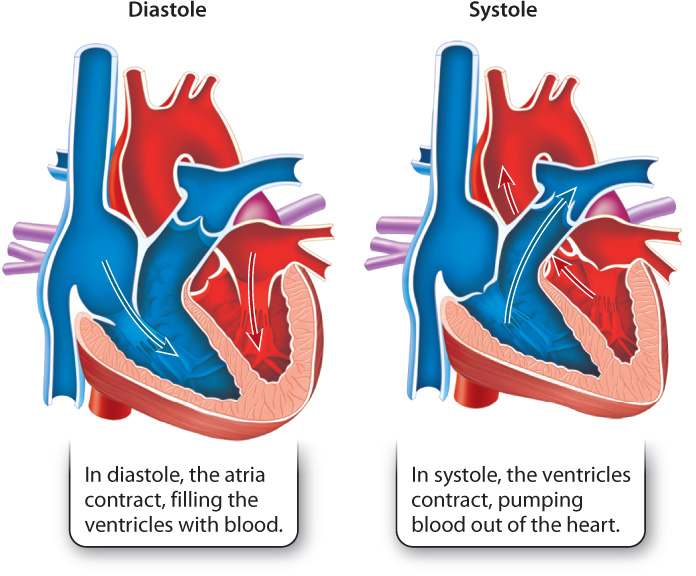
The contraction of the two atria followed by contraction of the two ventricles makes up the cardiac cycle (Fig. 39.22). The cardiac cycle is divided into two main phases. Systole is the contraction of the ventricles, and diastole is the relaxation of the ventricles. Systole, therefore, is the phase of the cardiac cycle in which blood is pumped from the heart into the pulmonary and systemic circulations, and diastole is the phase in which the atria contract and the ventricles fill with blood.
Quick Check 6
What pressure difference between the ventricle and the atrium causes the atrioventricular valve to close?
39.5.4 Cardiac muscle cells are electrically connected to contract in synchrony.
In order for the heart to function effectively as a pump, the cardiac muscle cells that make up the walls of the atria and ventricles must contract in a coordinated fashion. Atrial muscle cells must be activated to contract in synchrony during diastole to fill the ventricles, and ventricular muscle cells must contract in synchrony during systole to eject the blood from the heart. For cardiac muscle cells to pump in unison, they must be depolarized in unison by an action potential. In vertebrates, this action potential is transmitted by autonomic nerves that control the heart beat cycle.
Cardiac muscle cells have contractile properties similar to those of skeletal muscle cells (Chapter 37). However, cardiac muscle cells are distinct in two ways. First, specialized cardiac muscle cells can generate action potentials on their own, independently of the nervous system. Second, cardiac muscle cells are in electrical continuity with one another, meaning that they can pass their action potentials to adjacent cells. These two features of cardiac muscle cells ensure that all muscle cells in a region surrounding a heart chamber are activated and contract in unison. The specialized cardiac muscle cells capable of generating action potentials independently function as a pacemaker that causes the heart to beat with a basic rhythm. These cells stimulate neighboring cells to contract in synchrony. They are found in two specialized regions of the heart—the sinoatrial (SA) and atrioventricular (AV) nodes (Fig. 39.23).
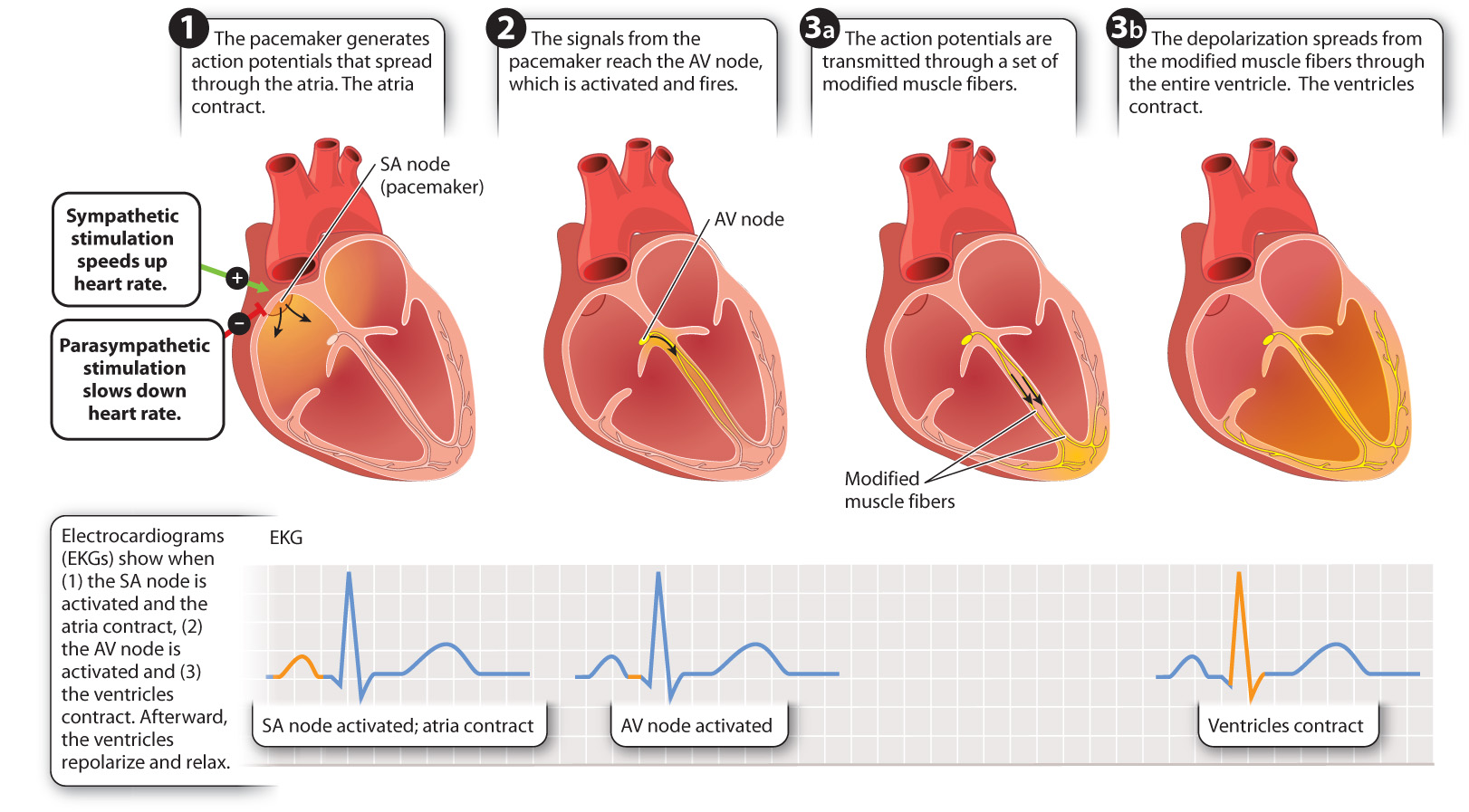
Whereas most nerve cells maintain their resting membrane potential until they receive a signal from another cell, the critical feature of pacemaker cells is that their resting membrane potential gradually becomes less negative on its own until it reaches threshold and the cell fires an action potential. The tendency of their membrane potential to become less negative is due to slow leakage of sodium ions into the cell and decreased flow of potassium out of the cell. When the cell reaches threshold, voltage-gated calcium ions open, causing more rapid depolarization. Like nerve cells, after firing an action potential, the pacemaker cells repolarize to reach their resting potential, keeping the heart relaxed between contractions. Also like nerve cells, pacemaker cells repolarize by opening voltage-gated potassium channels to allow potassium ions to leave the cell.
The heartbeat is initiated at the sinoatrial node, located at the junction of the vena cava and right atrium (Fig. 39.23). Because adjacent cardiac muscle cells are in electrical contact, action potentials initiated at the sinoatrial node spread rapidly from one cell to the next. Thus, when the pacemaker cells in the SA node fire an action potential, their depolarization spreads electrically throughout the right and left atria, causing them to contract in unison. Because there is no electrical contact between the atria and ventricles, the ventricles do not contract. Instead, the depolarization in the atria reaches a second set of pacemaker cells, located in the atrioventricular (AV) node.
Activation of the AV node transmits the action potential to the ventricles. A modified set of cardiac muscle fibers transmits the action potential from the AV node to the base of the ventricles. From there, the depolarization spreads throughout the ventricle walls, causing the ventricles to contract in unison. The delay in transmission from the AV node and through the conducting fibers ensures that the ventricles do not contract until they are fully filled with blood from the atria.
Electrodes placed over the surface of the chest and other body regions can record the electrical currents produced by the depolarization of the heart while it beats. This type of recording is called an electrocardiogram (EKG, because “kardia” is Greek for heart, although ECG is also used; Figure 39.23). Cardiologists use EKGs to diagnose heart problems or to determine what region of the heart has been damaged following a heart attack.
39.5.5 Cardiac output is regulated by the autonomic nervous system.
The volume of blood pumped by the heart over a given interval of time is its cardiac output (CO). This quantity is the key measure of heart function. Cardiac output is determined by calculating the product of heart rate (HR) and the volume of blood pumped during each beat, the stroke volume (SV): CO = HR × SV.
For example, if heart rate increases 50% and the heart’s stroke volume increases 10%, the heart’s cardiac output increases 65% ((1.5 × 1.1) − 1 × 100%). Cardiac output rises or falls in response to the metabolic demand for O2, which in turn depends on an animal’s state of activity. When an animal sleeps, cardiac output is minimal, but when it runs, cardiac output increases to meet the demand for O2 of its active muscles. Animals can increase their cardiac output by increasing heart rate or stroke volume or both. Whereas humans and other mammals rely most heavily on increases in heart rate to increase cardiac output, fish rely more on increases in stroke volume.
The nervous system controls the rate of the heartbeat. Nerve signals influence the rate at which the cardiac pacemaker cells depolarize in between action potentials. When there is no signal from the nervous system, the depolarization rate of the pacemaker cells sets the resting heart rate. Stimulation by sympathetic nerves causes the pacemaker cells of the sinoatrial node to depolarize more rapidly and the heartbeat to speed up. Stimulation by parasympathetic nerves causes these cells to depolarize more slowly and the heartbeat to slow down. By acting as a hormone (Chapter 38), adrenaline released by the adrenal gland into the circulation also elevates the heart rate for more prolonged periods of time. This hormone is a key component of the fight-or-flight response of many vertebrate animals.
Stroke volume is adjusted directly in response to changes in blood flow and heartbeat strength. It depends on how much blood returns to fill the heart and how strongly the heart contracts. These two processes are linked in order to match heart filling (that is, venous return) to heart emptying. During exercise, when an animal’s limb muscles regularly contract, their action increases the return of blood to the heart. The influx of additional blood stretches the walls of the atria and ventricles to a greater extent, causing them to contract more forcefully. As a result, more blood is ejected during each heartbeat.
The correspondence of stroke volume to changes in heart filling is described by Starling’s Law. It provides the means by which venous return and cardiac output are adjusted similarly to meet changing rates of blood supply. Increased sympathetic stimulation to the heart and circulating adrenaline also increase the contractile strength of the heart muscle, further increasing the heart’s stroke volume. These changes in heart rate and stroke volume underlie the increase in cardiac output that occurs when an animal changes from resting to activity (see Fig. 39.17).
In addition to the transport of respiratory gases, nutrients, and wastes, circulation is linked to several other bodily functions. The distribution of blood flow, controlled by changes in arterial resistance through vasodilation or vasoconstriction, is an important factor controlling heat loss or gain between an animal’s body and its environment. Thus, the circulatory system plays a central role in temperature regulation of many animals (Chapters 35 and 40). The circulatory system also provides the route by which hormones (Chapter 38) and immune cells (Chapter 43) are distributed throughout the body, enabling the endocrine system to maintain homeostatic control of diverse bodily functions and the immune system to defend the body against injurious microbes and toxic compounds. In doing so, the circulatory system serves as a general pathway by which hormonal and immune communication occurs to regulate, integrate, and protect the functional state of the whole organism.