40.1 PATTERNS OF ANIMAL METABOLISM
As discussed in Chapter 7, the biochemical pathways that make up animal metabolism are remarkably conserved among diverse groups of organisms. These pathways evolved early in the history of life and have been retained throughout the evolution and diversification of life. In Chapter 7, we examined the pathway of glucose breakdown by cellular respiration, which provides ATP to fuel the work of a cell. Here, we put this and related pathways in an organismal context. We explore how carbohydrates, proteins, and fats are metabolized for building body structures and supplying energy needs and how metabolic rate varies with an animal’s level of activity, internal body temperature, and size.
40.1.1 Animals rely on anaerobic and aerobic metabolism.
Nearly all animals depend on three main molecules as sources of energy and building blocks for growth and development: carbohydrates, fats, and proteins. The conversion of these food sources into biologically useful forms typically involves synthesizing adenosine triphosphate (ATP). While other nucleic acid triphosphates also serve as usable forms of energy, ATP is by far the most common one used by animals. The breakdown of carbohydrates, fats, and proteins to produce ATP involves a linked set of chemical reactions. Reactions that break down food sources to fuel the energy needs of a cell are referred to as catabolic. In contrast, reactions that result in net energy storage within cells and the organism are anabolic.
Let’s first review the breakdown of carbohydrates, including sugars (such as glucose) and starches, food sources that are chemically similar but clearly different in taste and texture. Glucose can be partially broken down in the absence of oxygen by glycolysis (Fig. 40.1). Glycolysis occurs in the cytosol of the cell and results in the production of two molecules of pyruvate and two molecules of ATP from the breakdown of each glucose molecule. If oxygen is not present, or is present only in small amounts, pyruvate is converted by fermentation into lactic acid. The production of ATP by anaerobic (“absence of oxygen”) metabolism provides rapid but short-term energy to the cell and organism.
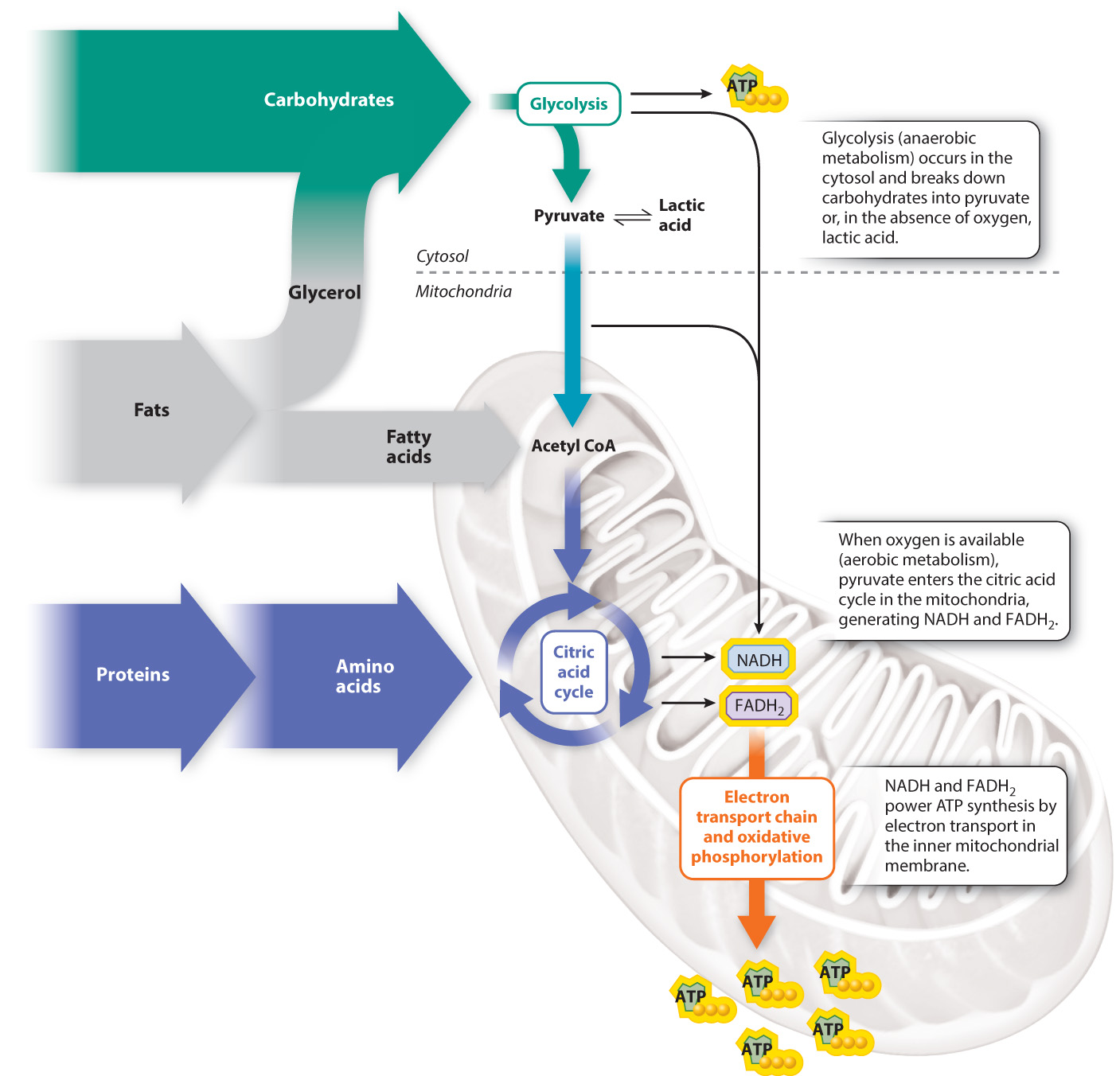
In contrast, aerobic (“of oxygen”) metabolism (specifically, cellular respiration) carried out within the mitochondria of eukaryotic animals provides a steady supply of ATP for longer-term, sustainable activity. When enough oxygen diffuses into the mitochondria, pyruvate can be processed by the citric acid cycle rather than converted to lactic acid (Fig. 40.1). In a series of oxidation–reduction reactions, energy-rich NADH and FADH2 molecules are produced. These molecules then enter the electron transport chain in the inner mitochondrial membrane. The electron transport chain couples the transfer of electrons to the pumping of protons across the inner mitochondrial membrane, resulting in a proton electrochemical gradient that enables the synthesis of ATP by oxidative phosphorylation. Because oxygen is reduced to water in the process, oxygen is consumed and water is produced.
Overall, aerobic respiration results in the formation of a total of 32 molecules of ATP from 1 molecule of glucose, much more than the 2 molecules of ATP from 1 molecule of glucose produced anaerobically. The metabolic efficiency, defined as the amount of energy captured in a usable form (approximately 286 kcal of useful ATP energy) per amount of energy in the starting molecule (686 kcal in one mole of glucose), is therefore approximately 42% (or 286/686 × 100). The remaining 58% of this energy is converted to heat and either lost or used to warm the animal. Glucose that is not metabolized is stored in the form of glycogen, primarily in the liver and muscles.
Lipids are another important energy source for most animals. Lipids, such as triacylglycerol, consumed in the diet are broken down to glycerol and free fatty acids, which enter glycolysis or the citric acid cycle to yield ATP by mitochondrial electron transport (Chapter 7). Alternatively, they can be stored as fat (adipose tissue), a particularly efficient storage form of energy. This is because fat can be stored in a form that yields more than twice the energy supply per unit weight (9.4 kcal/g) as glycogen (4.2 kcal/g). Fats are broken down into glycerol and free fatty acids, which enter glycolysis or the citric acid cycle to yield ATP by mitochondrial electron transport.
Proteins consumed in the diet are also a useful energy source for animals. Carnivores such as cats get most of their energy from the protein in the meat that they eat. Proteins, which are needed for building and maintaining the body, constitute the enzymes and structural elements of cells and tissues. Only following prolonged food deprivation, when fat and carbohydrate reserves are depleted, do animals break down protein reserves (mainly from their muscles) to form ATP. Animals preferentially rely on fats for long-term energy supply. Well-nourished humans have enough fat reserves to support their metabolic needs for 5–6 days before they must rely on protein reserves. Because nerve cells depend primarily on glucose to produce ATP, carbohydrates are spared for the brain and nervous system when animals face starvation.
Quick Check 1
Muscle protein, fat, and glycogen are all reservoirs of energy. In what order are they used during a prolonged fast?
40.1.2 Metabolic rate varies with activity level.
An animal’s overall rate of energy use defines its metabolic rate. Metabolic rate can be measured by the animal’s rate of oxygen consumption, which in turn reflects the aerobic production of ATP (Fig. 40.2). In 1773, Antoine Lavoisier first demonstrated that animal respiration is the same as combustion, as observed in the burning of a candle. A burning candle placed inside a sealed container is eventually extinguished. Similarly, a bird placed inside a sealed container eventually loses consciousness. When the candle and the bird were placed in the chamber together, both succumbed more quickly. Borrowing from Joseph Priestley’s discovery of oxygen as the gas in air that reacts with metals (in the process now called oxidation), Lavoisier recognized that both the bird and candle consumed oxygen, or “pure air” as he called it, converting it into “fixed air” (carbon dioxide), or air of “poorer quality.”
Metabolic rate is affected by many factors, one of which is the activity level of an organism. When an animal shifts from rest to activity, its metabolic rate and oxygen consumption rise to meet its increased demand for ATP. The onset of activity requires immediate energy, which in animals is provided by specialized energy stores in their tissues. In vertebrate muscle cells, for example, phosphocreatine, a ready source of high-energy phosphates, is hydrolyzed to synthesize ATP directly from ADP. This reaction is followed by the relatively rapid production of ATP by anaerobic glycolysis. Although glycolysis produces relatively few ATP molecules per molecule of glucose broken down, the reactions are extremely fast, providing a rapid short-term supply of energy for animals. Animals rely on anaerobic glycolysis for short bursts of intensive activity.
Eventually, enough oxygen diffuses into the mitochondria to allow aerobic respiration to occur at a rate that meets the ATP needs of the animal. Therefore, the rate of oxygen consumption initially increases, then levels off (Fig. 40.3). At this point, the animal’s need for energy is being met entirely by aerobic respiration.
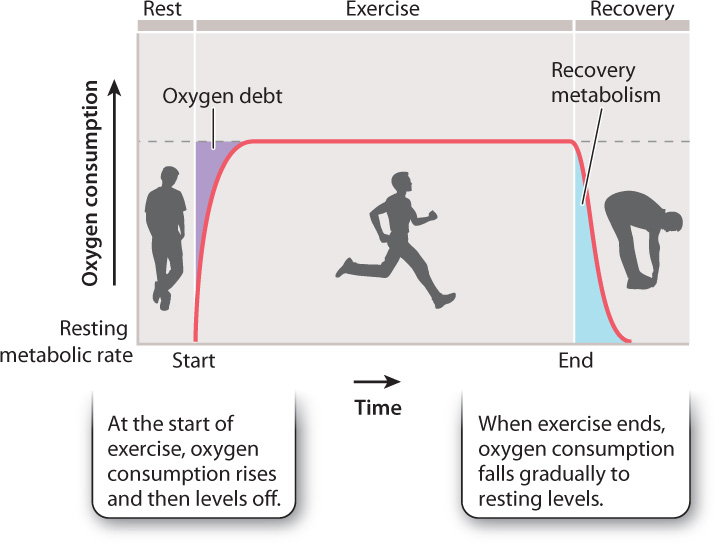
With even more prolonged or intense activity, more ATP is needed, and it is harder to meet that need by aerobic metabolism alone. Your experience tells you that you can walk at a comfortable pace for a long period of time, but when you run, you tire more quickly. And the faster you run, the more quickly you tire. This is because more intensive physical exercise requires a greater reliance on anaerobic glycolysis to produce ATP. The resulting buildup of lactic acid and decrease in pH inside the muscle cells force an animal to decrease its activity. The production of lactic acid through fermentation limits an animal’s performance by lowering the pH of the animal’s blood, producing a condition called metabolic acidosis. Metabolic acidosis contributes to fatigue.
Sprinters rely heavily on anaerobic ATP production for intense short-term bouts of activity, but longer races are run at progressively slower speeds for increased endurance as the runner relies more heavily on aerobic metabolism to produce ATP. The world record for the 100-m sprint averages just above 10 m/s, but for the 1500-m race it is about 6.6 m/s.
When activity ends, the animal’s oxygen consumption rate declines but does not immediately return to resting levels, as shown in Fig. 40.3. The elevated consumption of oxygen following activity is the animal’s recovery metabolism. It represents the continued metabolic energy required to reestablish the resting metabolic state of the cells. During recovery metabolism, cells re-synthesize depleted ATP stores and metabolize the end products of fermentation, particularly lactic acid. The difference between an animal’s immediate energy need at the onset of activity versus energy supplied by aerobic metabolism is often referred to as the animal’s “oxygen debt.” This debt is “paid back” following exercise by the animal’s recovery metabolism. Recovery metabolism is associated with elevated breathing and heart rates that you (and other animals) experience when resting after moderate to intense exercise.
Case 7 Predator–Prey: A Game of Life and Death
40.1.3 Does body temperature limit activity level in predators and prey?
We just saw that metabolic rate increases with activity level. A by-product of metabolism is the generation of heat, which is a necessary consequence of the second law of thermodynamics (Chapters 1 and 6). Animals have several mechanisms for dissipating excess heat, including sweating, changes in blood flow, and (in dogs, for example) panting.
Cheetahs, as top predators, are one of the fastest land animals known today, capable of speeds up to 70 mph (114 km/h). Such extreme speed generates a lot of heat. Interestingly, instead of releasing the excess heat as it builds up, the cheetah stores it, dissipating it only after a chase.
The amount of heat that cheetahs store during a sprint can be considerable, on the order of 60 times the heat produced at rest. Heat storage raises the cheetah’s body temperature. Cheetahs can therefore reach remarkable speeds but can sustain them only for short durations and must rest for long periods of time between sprints. Because their bodies can support only so much heat storage, cheetahs do not run when the temperature of their environments reaches approximately 105°F (41°C). In other words, the amount of time a cheetah runs and the distance it covers are limited by its body temperature.
Gazelles, common prey of cheetahs, have similar heat storage mechanisms and are similarly known for their speed and agility. Goats, by contrast, do not store heat as they increase their activity levels, instead dissipating it through evaporative mechanisms. But goats are not known for their speed. Heat storage therefore seems to enable quick bursts of speed, and has evolved as an adaptation among both predators and prey to achieve short, rapid sprints.
40.1.4 Metabolic rate is affected by body size.
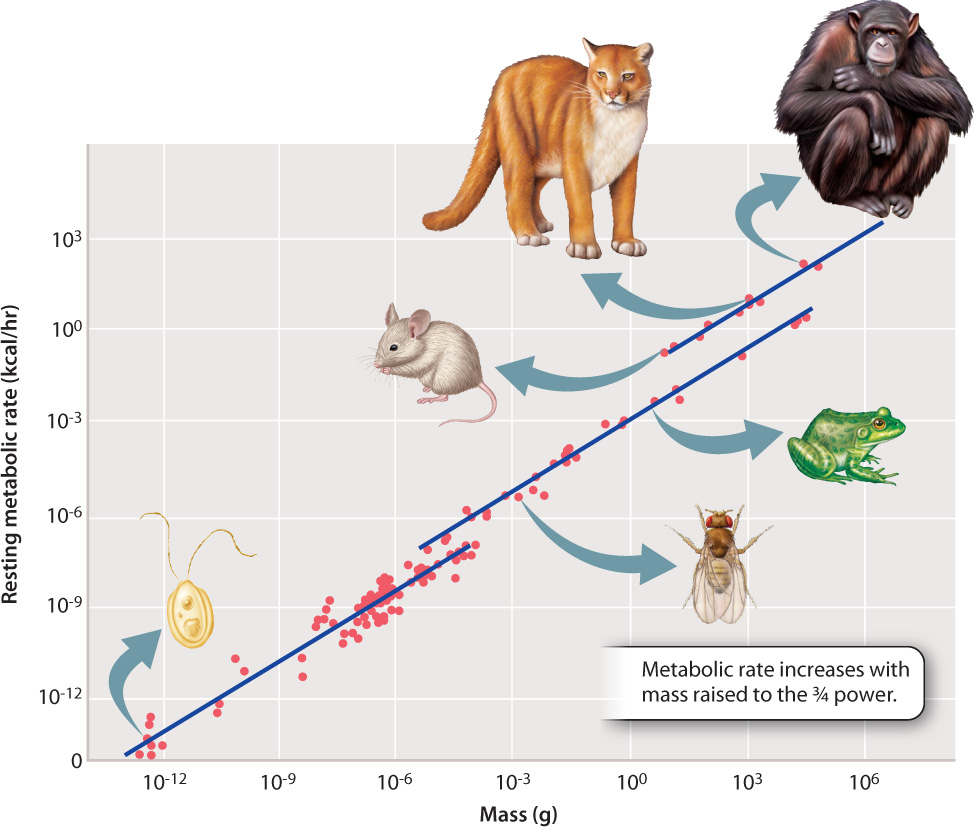
In addition to activity level, an animal’s size influences its metabolic rate. At rest, larger animals consume more energy and have higher metabolic rates than smaller ones. However, resting, or basal, metabolic rate does not increase linearly with an animal’s mass. Instead, measurements of resting metabolic rate in a wide range of organisms show that metabolic rate increases with animal mass raised to the 3/4 power (Fig. 40.4). This means that the average rate at which each gram of body tissue consumes energy is less in larger animals compared to smaller ones. Or, put another way, the larger the organism, the lower the metabolic rate per gram of body tissue. This scaling pattern of cellular energy metabolism is remarkable because it holds across a diverse size range of unicellular and multicellular organisms, as can be seen in Fig. 40.4.
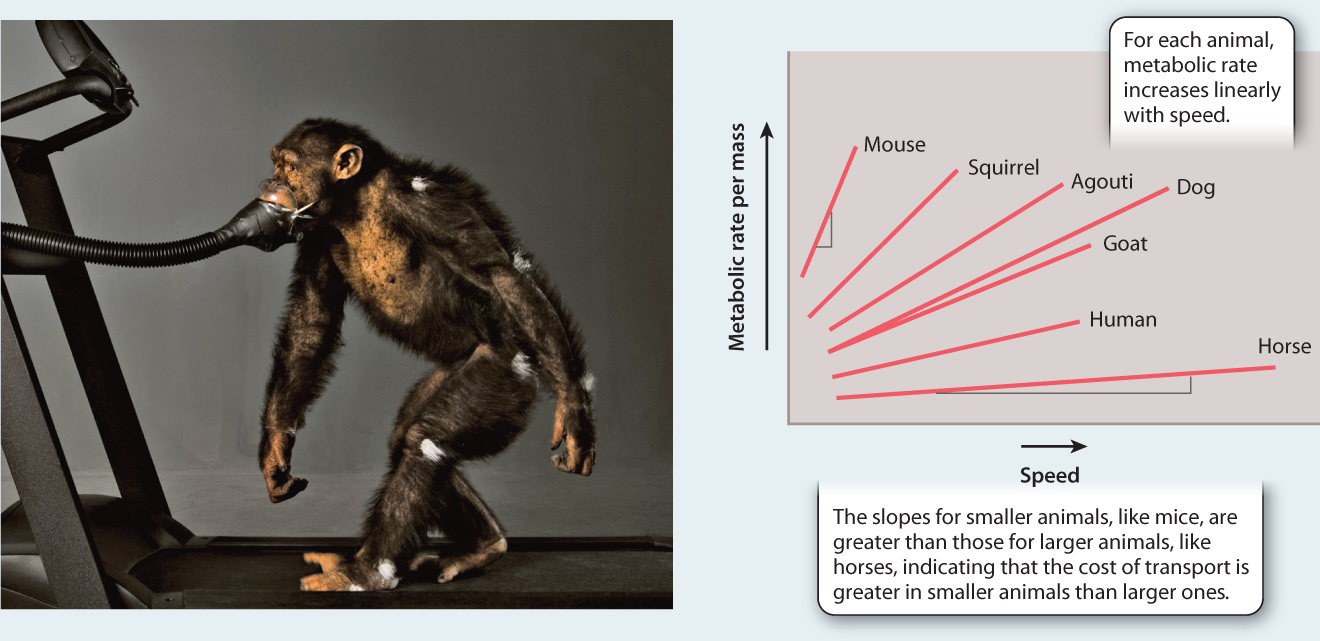
In addition to basal metabolic rate, we can also examine how metabolic rate varies with respect to exercise in different-sized animals. In this case, for example, we are looking at how energy is used to allow animals to move at different speeds. The American comparative physiologist C. Richard Taylor has extensively studied how metabolic rate varies with running speed in different-sized terrestrial animals (Fig. 40.5). He found that metabolic rate increases linearly with speed, and that larger animals expend less energy per unit mass than smaller ones.
FIG. 40.5How is metabolic rate affected by running speed and body size?
BACKGROUND In the 1970s and 1980s, the American physiologist C. Richard Taylor and his colleagues performed studies on the relationship between metabolic rate and running speed in mammals of different sizes. They were interested in understanding the energetic costs of running in different animals.
EXPERIMENT Taylor and colleagues measured oxygen consumption during running for a wide range of organisms. For each, they trained the animals to run on a treadmill at different speeds and measured oxygen consumption using either a face mask or enclosure. Oxygen consumption was used as a measure of metabolic rate.
RESULTS The researchers found that there is a linear increase in metabolic rate with speed in different-sized animals and that larger animals expend less energy per unit body mass to move a given distance compared to smaller ones, as shown on the graph below.
FOLLOW-UP WORK Similar studies were performed with kangaroos, which move by hopping rather than running. Interestingly, it was found that it is “cheaper” (that is, it requires less energy) to hop than to run. This finding perhaps explains why kangaroos and other hopping animals survived, while many animals that run on four legs became extinct in Australia 20,000 to 30,000 years ago with the arrival of early humans. Studies have also compared the energetic costs of running on two legs and running on four legs, with the finding that the cost is the same.
SOURCES Dawson, T. J., and C. R. Taylor. 1973. “Energetic Cost of Locomotion in Kangaroos.” Nature 246 : 313–314; Taylor, C. R., N. C. Heglund, and G. M. O. Maloiy. 1982. “Energetics and Mechanics of Terrestrial Locomotion.” Journal of Experimental Biology 97:1–21.
Quick Check 2
Does a dog that is twice as heavy as a cat have twice the resting metabolic rate?
40.1.5 Metabolic rate is linked to body temperature.
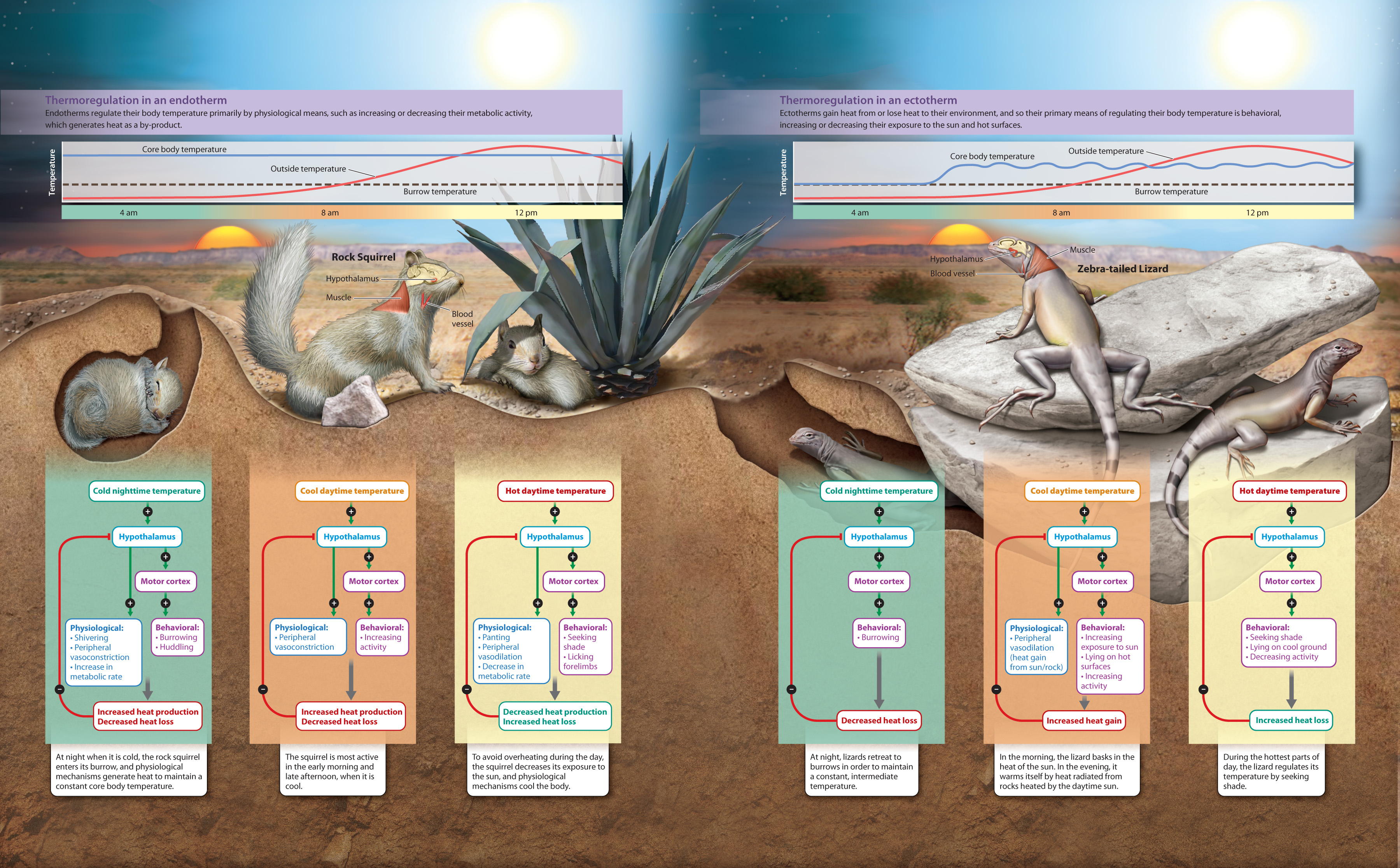
Metabolic rate is also affected by an animal’s internal body temperature. Temperature affects the rates of chemical reactions, which in turn determine how fast fuel molecules can be mobilized and broken down to supply energy for a cell. Animals can be categorized by the sources of most of their heat. Animals that produce most of their own heat as by-products of metabolic reactions, including the breakdown of food, are endotherms. These animals usually, but not always, maintain a constant body temperature that is higher than that of their environment. As we saw earlier (Chapter 35), the maintenance of a constant core body temperature is a form of homeostasis and requires balancing heat production and heat loss. Mammals, including humans, and birds are endotherms.
By contrast, animals that obtain most of their heat from the environment are ectotherms. Ectotherms often regulate their body temperatures by behavioral means such as moving into or out of the sun. Think of a turtle or another reptile sunning itself on rock: It is raising its body temperature. The temperatures of these animals usually fluctuate with the outside environment. Most fish, amphibians, reptiles, and invertebrates are ectotherms.
Endotherms are sometimes referred to as “warm-blooded” and ectotherms as “cold-blooded,” but these terms are misleading because, depending on environmental conditions, cold-blooded animals can have core body temperatures higher than warm-blooded animals. It should also be kept in mind that these thermoregulatory mechanisms are not completely distinct, but instead represent extremes of a continuum.
Nevertheless, thermoregulatory mechanisms have profound implications for an animal’s metabolic rate. Endotherms have a higher metabolic rate than ectotherms. As a result, they are able to be active over a broader range of external temperatures than ectotherms. The activity level and metabolic rate of ectotherms both increase with increasing body temperature. However, ectotherms have metabolic rates that are about 25% of endotherms of similar body mass and at similar body temperatures. Although they can achieve activity levels similar to those of endotherms when their body temperatures are similar, ectotherms cannot sustain prolonged activity. Therefore, ectotherms such as lizards or insects rely on brief bouts of activity, followed by longer periods of inactivity compared to endotherms.
Endotherms benefit from having an active lifestyle. However, their high metabolic rate requires long periods of foraging to acquire enough food to keep warm and be active. By contrast, although ectotherms are limited in their ability to sustain activity, they can survive much longer periods without food.
As we have discussed, the control of core body temperature, or thermoregulation, is a form of homeostasis and requires the coordinated activities of the nervous, muscular, endocrine, circulatory, and digestive systems. We summarize how these systems work together in endotherms and ectotherms in Fig. 40.6.
Visual Synthesis: Homeostasis and Thermoregulation
Click the image below to view an enlarged version in a new window.