42.1 THE EVOLUTIONARY HISTORY OF REPRODUCTION
While the ability to reproduce is shared across all domains of life, the particular way in which animals reproduce varies among species, from the familiar to what might strike us as truly bizarre. To take just one of many examples: After birth, a male anglerfish finds a female anglerfish, bites her skin, and fuses with her. His body atrophies, leaving the male reproductive organs physically attached to the female and allowing her eggs to be fertilized when she is ready. To begin to make sense of the diversity of reproductive strategies, we consider how different forms of reproduction evolved, what organisms use them, and their adaptive significance.
42.1.1 Asexual reproduction produces clones.
Organisms have two potential ways to reproduce. One is the production of genetically identical cells or individuals called clones. This form of reproduction is called asexual reproduction, and it occurs in a wide variety of organisms (Fig. 42.1).
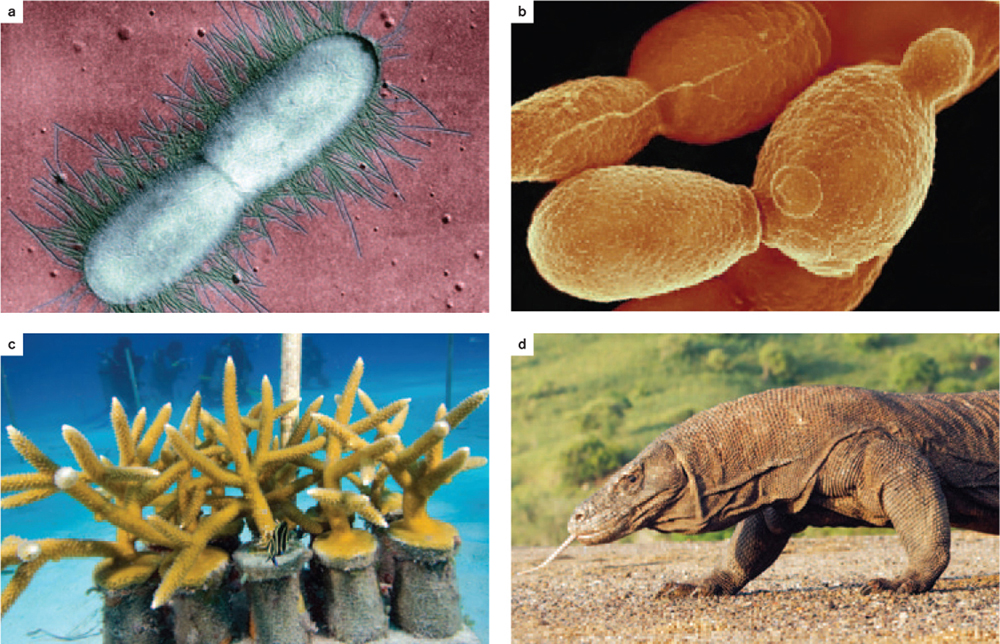
For example, bacteria (Fig. 42.1a) and archaeons reproduce by binary fission, a form of cell division in which the genome replicates and then the cell divides in two (Chapter 11). The result is two cells that are either identical to the parent cell or nearly so, differing only because of chance mutations.
Prokaryotes can nevertheless increase genetic variation in various ways (Chapter 26). For example, they can transfer DNA from one individual to another, usually of the same species, and much more rarely to an individual of another species, during conjugation. They can also obtain DNA directly from the environment and by viral infection, incorporating genes that might have evolved in even distantly related species. These processes allow prokaryotes to acquire novel genetic sequences that can affect fitness.
Eukaryotic cells divide by mitosis (Chapter 11), a process that evolved early and was present in the common ancestor of all living eukaryotes. Mitosis is a form of growth and development in all eukaryotes, and is the basis of asexual reproduction in many eukaryotes.
One form of asexual reproduction in fungi, plants, and some animals is budding, in which a bud, or protrusion, forms on an organism and eventually breaks off to form a new organism that is smaller than its parent. The budding yeast, Saccharomyces cerevisiae, which is used to make bread and beer, is so named because of its ability to produce new individuals by budding (Fig. 42.1b). Budding can also occur in animals such as the water-dwelling hydra, which can grow and subsequently break off part of itself to form a new, free-living individual. Whether used to describe reproduction in unicellular or multicellular organisms, budding involves an unequal division between a mother and daughter cell or organism. Budding is the result of mitosis and therefore, like binary fission, results in an offspring that is genetically identical to the parent.
Another form of asexual reproduction is fragmentation, in which new individuals arise asexually by the splitting of one organism into pieces, each of which develops into a new individual. Certain molds, algae, worms, sea stars, and corals reproduce by fragmentation. While fragmentation occurs naturally, it is also done intentionally to propagate corals in reef restoration projects (Fig. 42.1c). Once again, the new individual is a clone of the parent.
A final example is parthenogenesis, literally “virgin birth.” In this form of asexual reproduction, females produce eggs that are not fertilized by males but divide by mitosis and develop into new individuals. Parthenogenesis occurs in invertebrates, such as bees, other insects, and crustaceans, as well as in vertebrates, including some fish and reptiles (Fig. 42.1d). The details of parthenogenesis differ depending on the organism. Egg cells are usually the product of meiosis and are haploid, having one set of chromosomes. In some organisms, this haploid egg cell divides by mitosis to become a haploid adult; in others, there is a doubling of the DNA content during development, so the adult is diploid, having two sets of chromosomes; and in still others, the egg cell does not undergo meiosis (it is diploid) and divides by mitosis to become a diploid adult. In all cases, parthenogenesis involves a single maternal parent.
42.1.2 Sexual reproduction involves the formation and fusion of gametes.
The second way that organisms reproduce is by combining a complete set of genetic information from two individuals to make a new genetically unique individual. This form of reproduction is called sexual reproduction.
At its core are two basic biological processes—meiosis and fertilization (Fig. 42.2). Meiosis is a form of cell division that halves the number of chromosomes; fertilization involves fusion of specialized cells to restore the original chromosomal content. Meiosis evolved in the ancestors of modern eukaryotes, and so sexual reproduction occurs only in eukaryotes.
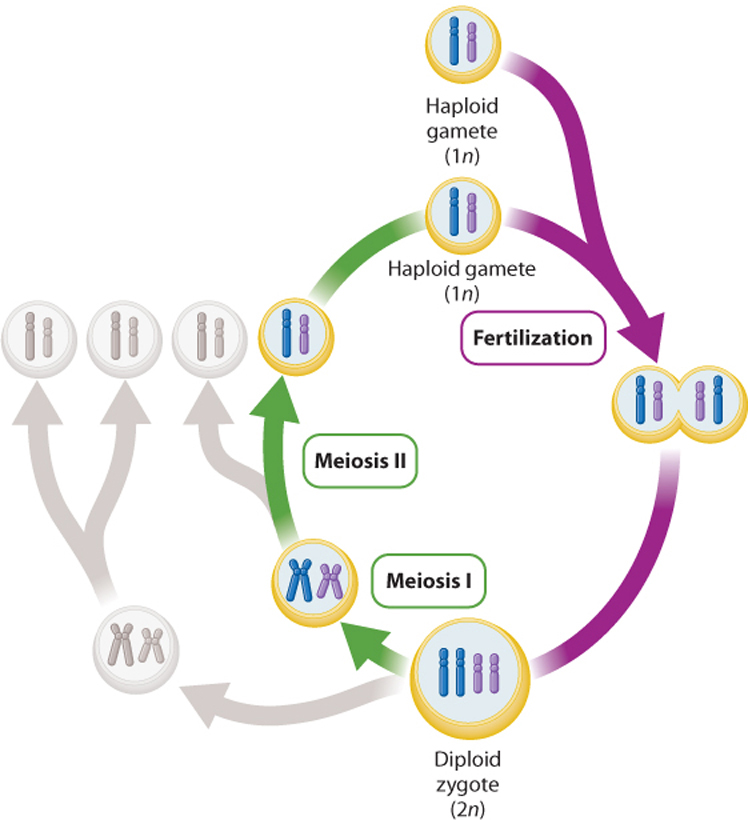
As we saw in Chapter 11, meiosis consists of one round of chromosome replication followed by two rounds of cell division, resulting in four cells each with half the number of chromosomes of the parent cell. So, in a diploid species with 2n chromosomes, the result of meiosis is four cells with n chromosomes. The resulting cells are haploid and are called gametes or spores. Humans, for example, have 23 pairs of chromosomes (2n = 46 chromosomes) and human gametes have half this number (n = 23 chromosomes).
Many species produce two types of gametes that differ in shape and size. The smaller, male gametes are called spermatozoa, or sperm, and the larger, female gametes are called ova, or eggs. For other organisms, including green algae, diatoms, slime molds, and most fungi, the two types of gametes are the same size. In spite of their similar size and appearance, however, they come in two (or more) distinct mating types and cannot reproduce sexually with individuals of the same type.
Fertilization involves the fusion of two gametes, one from each parent (Fig. 42.2). The result is a zygote. If each gamete has n chromosomes, the resulting zygote has 2n chromosomes. Fertilization results in a net reduction in cell number—two haploid gametes fuse to make one diploid zygote. For unicellular organisms with a prominent haploid phase, increase in population size occurs when the zygote undergoes meiosis and the four resulting cells divide by mitosis (Chapter 27). For diploid multicellular organisms, however, fertilization is the direct basis for reproduction. The zygote divides by mitosis and develops into an embryo, an early stage of multicellular development.
Meiosis and fertilization have significant genetic consequences. During meiosis, pairs of homologous chromosomes undergo recombination, shuffling the combination of alleles between the chromosomes. In addition, meiosis randomly sorts homologous chromosomes into different gametes. As a result, the products of meiosis, the haploid gametes, are each genetically unique and represent a mixture of the parental genetic makeup. Then, during fertilization, two of these unique haploid gametes fuse, creating new genetic combinations and a unique diploid individual.
Quick Check 1
Sexual reproduction results in offspring that are genetically different from one another and from their parents. Describe four mechanisms that produce this genetic variation. Are any of these mechanisms shared with asexual reproduction?
42.1.3 Many species reproduce both sexually and asexually.
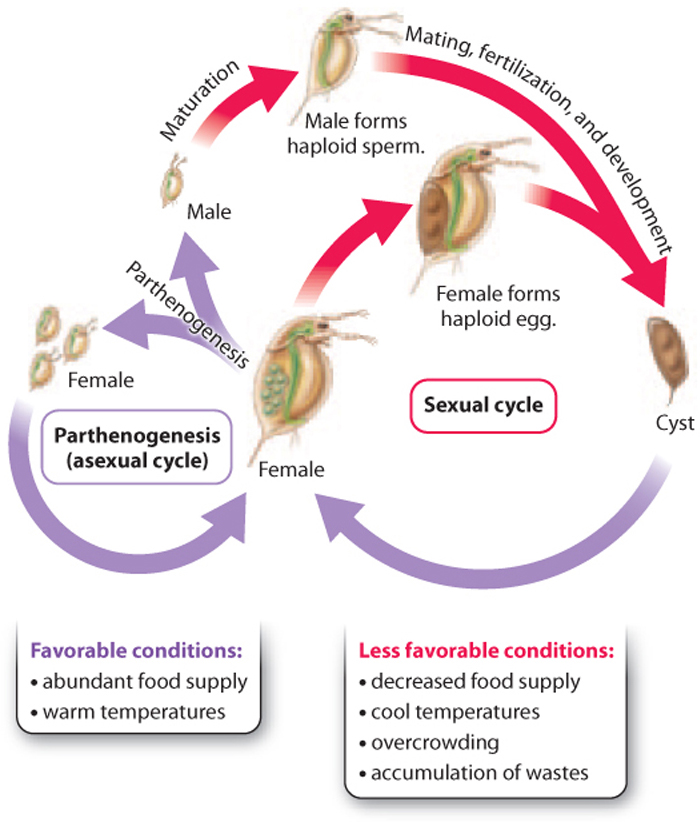
Most organisms that reproduce asexually are also capable of reproducing sexually. For example, hydras reproduce asexually by budding, but this ability to reproduce asexually does not preclude their ability to reproduce sexually. Even budding yeast are capable of sexual reproduction. Most aphids, fish, and reptiles capable of parthenogenesis can also reproduce sexually.
What determines when an organism reproduces sexually and when it reproduces asexually? This question has been examined in the freshwater crustacean Daphnia. Daphnia reproduce asexually by parthenogenesis in the spring, when food is plentiful, perhaps because asexual reproduction is a rapid form of reproduction that allows the organism to take advantage of abundant resources (Fig. 42.3). Later in the season, when conditions are less favorable because of increased crowding and decreased food and temperature, Daphnia reproduce sexually, mating to produce a zygote that starts to form an embryo. At an early stage, however, development is arrested and metabolism slows dramatically. The embryo forms a thick-walled structure, called a cyst, which enables it to resist freezing and drying and therefore last through the winter.
This link between environmental conditions and mode of reproduction has been observed in many other species, from algae to slime molds. The life cycle of Daphnia hints that asexual and sexual reproduction have costs and benefits, a subject we turn to now.
42.1.4 Exclusive asexuality is often an evolutionary dead end.
Asexual reproduction has some advantages compared to sexual reproduction. It does not involve finding and attracting a mate, which takes time and energy. Certain birds, for example, go to elaborate lengths to attract mates. Male birds of paradise invest a significant amount of energy in growing colorful plumage, and male bowerbirds take time to build elaborate homes for their potential mates (Fig. 42.4). For other organisms, finding a mate can be difficult because of very low population density—this is the case for anglerfish, and may help to explain their apparently unusual reproductive strategy, described earlier.

In addition, asexual reproduction is rapid, allowing organisms to increase their numbers quickly. Consider rates of population growth for organisms that reproduce asexually and those that reproduce sexually. In asexual organisms, all offspring can produce more offspring, allowing for exponential population growth if conditions are favorable. By contrast, in sexually reproducing organisms, only females produce new offspring. The consequence of this difference is that an asexual group can have a much greater rate of population growth compared to that of a sexually reproducing one. The British evolutionary biologist John Maynard Smith called this effect the twofold cost of sex (Fig. 42.5), and it is one of the key disadvantages of sexual reproduction.
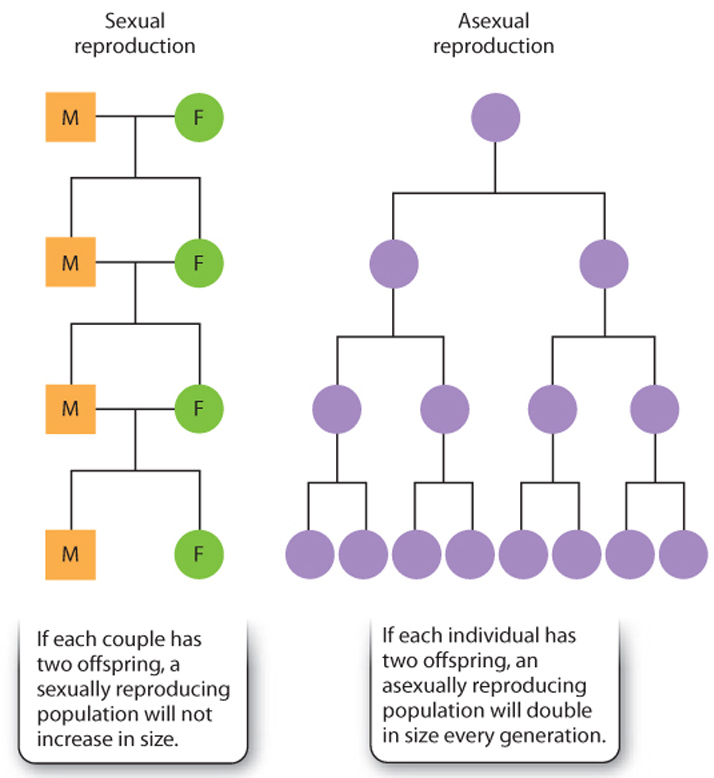
In spite of the advantages of asexual reproduction, a survey of the natural world reveals a striking pattern: Most organisms reproduce sexually at least part of the time. And even prokaryotes, as we have seen, have mechanisms for genetic exchange. The observation that the vast majority of organisms reproduce sexually at some point suggests that sexual reproduction offers advantages over asexual reproduction. Asexual reproduction is quick, but it produces offspring that are genetically identical to one another and to the parent. The only source of genetic variation is chance mutations. By contrast, one of the defining features of sexual reproduction is the production of offspring that are genetically different from one another and from their parents.
Many hypotheses have been developed to explain the advantages of producing genetically distinct offspring in spite of the costs of sexual reproduction. One suggests that sexual reproduction allows organisms to adapt faster than asexual ones because rare beneficial mutations that arise in different organisms can be brought together, increasing the overall fitness of the population. Another set of hypotheses suggests that sexual reproduction allows a population to purge itself of harmful mutations more quickly than could a population of asexual individuals.
A third hypothesis—the Red Queen hypothesis—suggests that sexual reproduction is a mechanism of parasite defense. This hypothesis takes its name from Lewis Carroll’s Through the Looking-Glass, in which the Red Queen tells Alice that she needs to keep running to stay in the same place. Similarly, in host–parasite coevolution, the host evolves defenses against the parasite, and the parasite evolves new ways to infect the host, so each evolves rapidly, or keeps moving, simply to maintain its same ecological position—that is, to stay in the same place. This kind of rapid evolution requires lots of genetic variation on which natural selection can act, and sexual reproduction increases genetic variation among individuals more than asexual reproduction does.
These different explanations for the prevalence of sexual reproduction are not mutually exclusive. It may be that several mechanisms work together, or that different ones are important for different organisms.
It is instructive to consider groups of eukaryotes that have lost the ability to reproduce sexually, and those that have lost the ability to reproduce asexually. While there are organisms such as the New Mexico whiptail lizard (Cnemidophorus neomexicanus) that reproduce only asexually, these species are not very old in evolutionary terms. In other words, there are very few, if any, ancient asexual groups. It appears that asexual reproduction on its own rarely allows a species to persist for very long in evolutionary terms.
A possible exception to this rule may be bdelloid rotifers, a group of microscopic freshwater invertebrates that are thought to have reproduced asexually for at least 35 to 40 million years (Fig. 42.6). The biology of this group is being intensely scrutinized for hints of its pattern of reproduction over time. If indeed bdelloid rotifers have persisted for tens of millions of years without sexual reproduction, then we have an interesting puzzle. We might predict that the rotifers have either an unusual way to escape the long-term evolutionary costs of genetic uniformity, or an as yet unknown mechanism for exchange of genetic material.
FIG. 42.6Do bdelloid rotifers only reproduce asexually?
BACKGROUND Many groups of animals reproduce asexually, but most of these organisms reproduce sexually at least some of the time. The bdelloid rotifer, one of several species of rotifer, may be an exception.
OBSERVATION All bdelloid rotifers found to date are females. No males or hermaphrodites (organisms with both male and female reproductive organs) have ever been observed. Meiosis has never been documented.
HYPOTHESIS Bdelloid rotifers are asexual and reproduce solely by parthenogenesis, in which unfertilized eggs develop into adult females.
PREDICTION In the absence of meiosis, the two copies (alleles) of a gene do not recombine and mutations that accumulate are not reshuffled between them. As a result, two alleles of a gene in an asexually reproducing organism will show a much higher degree of sequence difference compared to two alleles of a gene in a sexually reproducing organism.
RESULTS Following is the sequence from the two copies of the hsp82 gene in a sexually reproducing rotifer (Fig. 42.6a) and an asexually reproducing rotifer (Fig. 42.6b). As can be seen, the two copies are similar in Fig. 42.6a but very different in Fig. 42.6b.

CONCLUSION All evidence to date suggests that bdelloid rotifers are exclusively asexual.
SOURCE Welch, M. D., and M. S. Meselson, 2000. “Evidence for the Evolution of Bdelloid Rotifers Without Sexual Reproduction or Genetic Exchange.” Science 288 (5469):1211–1215.
Mammals are unusual in the opposite way. Mammals have lost the ability to reproduce asexually. Parthenogenesis, as we have seen, occurs in some vertebrates, but does not occur in mammals. Although this form of reproduction may seem odd from our point of view, scientists have wondered just the opposite—why can’t female mammals reproduce without a father? Asked another way, why do mammalian embryos need both a maternal and a paternal genome? Wouldn’t two maternal genomes from a single parent do the trick? Evidently not. Scientists have known for some time that mammalian maternal and paternal genomes are not quite the same. There are some genes, called imprinted genes, that are expressed differently depending on their parent of origin. Imprinting ensures that mammals develop from embryos with one maternal and one paternal genome. When researchers manipulated key imprinted genes in a maternal genome, they were able to breed a parthenogenetic mouse—one with two copies of a maternal genome from a single parent.
We can speculate that early mammals gained fitness advantages in the evolution of imprinting of genes. However, a consequence was the loss of parthenogenesis in mammals. Parthenogenesis might offer greater fitness advantages than strictly sexual reproduction in some species of mammals living today, but that option is no longer available.