42.4 GAMETE FORMATION TO BIRTH IN HUMANS
With an understanding of male and female reproductive anatomy and endocrinology, we are ready to follow human reproduction and development, from the formation of sperm and eggs, to fertilization itself, to subsequent development of the embryo, and finally to birth. Along the way, we consider how these processes vary in different organisms. Our understanding of these processes is in large part due to studies in model organisms, including the sea urchin (Strongylocentrotus purpuratus), roundworm (Caenorhabditis elegans), fruit fly (Drosophila melanogaster), African clawed frog (Xenopus laevis), and house mouse (Mus musculus). These organisms have particular reproductive and developmental features that make them amenable to laboratory study.
42.4.1 Male and female gametogenesis have shared and distinct features.
The formation of gametes is called gametogenesis. Gametogenesis follows the same steps in males and females, but there are striking differences in timing (Fig. 42.19). Spermatogenesis is the formation of sperm. It occurs in the seminiferous tubules of the testes, begins at puberty, and continues throughout life. The process starts with spermatogonia, diploid stem cells that divide by mitosis to ensure a continuous population of precursor cells. These cells differentiate into diploid primary spermatocytes, which undergo the first meiotic division to produce two secondary spermatocytes and the second meiotic division to produce four haploid spermatids joined by cytoplasmic bridges. They subsequently separate from each other to become distinct haploid cells. The maturation process, in which spermatids become spermatozoa or sperm, begins in the testes and continues in the epididymis, where they acquire motility. Spermatogenesis takes about 2 to 3 months.
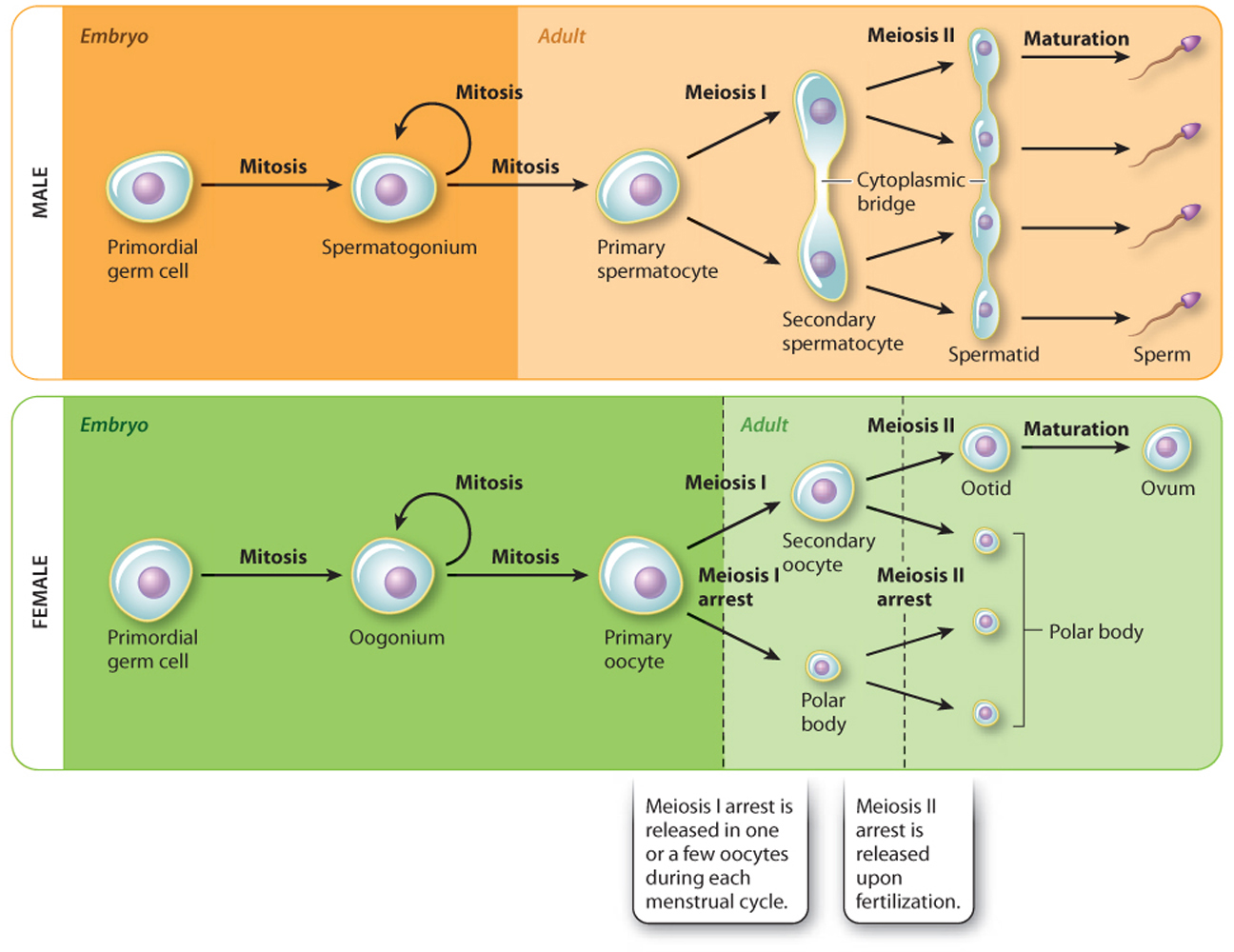
Oogenesis is the formation of ova or eggs. The diploid precursor cells are called oogonia. These cells form during embryonic development and divide by mitosis. Some oogonia differentiate into diploid primary oocytes during fetal development. These enter the first meiotic division but arrest immediately in prophase I, the first stage of meiosis. All these steps take place before the female is even born. At the time of birth, a female has about 1 to 2 million primary oocytes, all arrested in prophase I of meiosis. They do not resume the first meiotic division until a menstrual cycle. Therefore, remarkably, they all remain arrested for at least 12 years (from birth to the age of menarche) and some as long as 50 years (to the age of menopause).
In the first phase of the menstrual cycle, approximately 10 to 20 follicles begin to mature because of rising FSH levels. One of these follicles becomes dominant, allowing the primary oocyte to complete the first meiotic division. This division is asymmetric, leading to the formation of a large secondary oocyte and a much smaller polar body. The unequal division allows most of the cytoplasm to be partitioned into the secondary oocyte. The secondary oocyte is released from the ovary and immediately enters the second meiotic division, but again arrests, this time in metaphase II. The secondary oocyte remains arrested in metaphase II until fertilization takes place. Upon fertilization, the second meiotic division continues, producing an ootid and another polar body, again allowing most of the cytoplasm to end up in the developing egg. The ootid quickly develops into a mature ovum or egg.
Quick Check 4
When are primary oocytes and spermatocytes produced, and how many gametes result from one diploid cell in females and males?
42.4.2 Fertilization occurs when a sperm fuses with an oocyte.
Fertilization involves the fusion of gametes, restoring the diploid chromosome content (Fig. 42.20). In humans, male and female gametes are usually brought together by sexual intercourse. Sexual arousal leads to a host of physiological changes, including erection of the penis and erection of the clitoris and lubrication of the vagina. The erect penis is inserted into the vagina until orgasm occurs. Orgasm is characterized by feelings of pleasure and contractions of the pelvic muscles. In males, it is usually accompanied by ejaculation, in which semen, containing sperm, is delivered into the vagina.
Sperm travel through the cervix and into the uterus and fallopian tubes. Fertilization usually occurs in one of the fallopian tubes. As a result, many of the details of how sperm travel through the female reproductive tract are poorly understood. Sperm are motile, but from studies of the time it takes sperm to reach the fallopian tubes, it appears that the female reproductive tract plays an active role in moving sperm to the oocyte, for example through muscular contractions.
In the female reproductive tract, sperm go through a key developmental step, called capacitation, a process that is unique to mammals. Capacitation is a series of physiological changes that allow the sperm to fertilize the egg. These changes include alterations in the fluidity of the plasma membrane, loss of some surface membrane proteins, and changes in the charge across the membrane, called the membrane potential. As a result, sperm show increased motility.
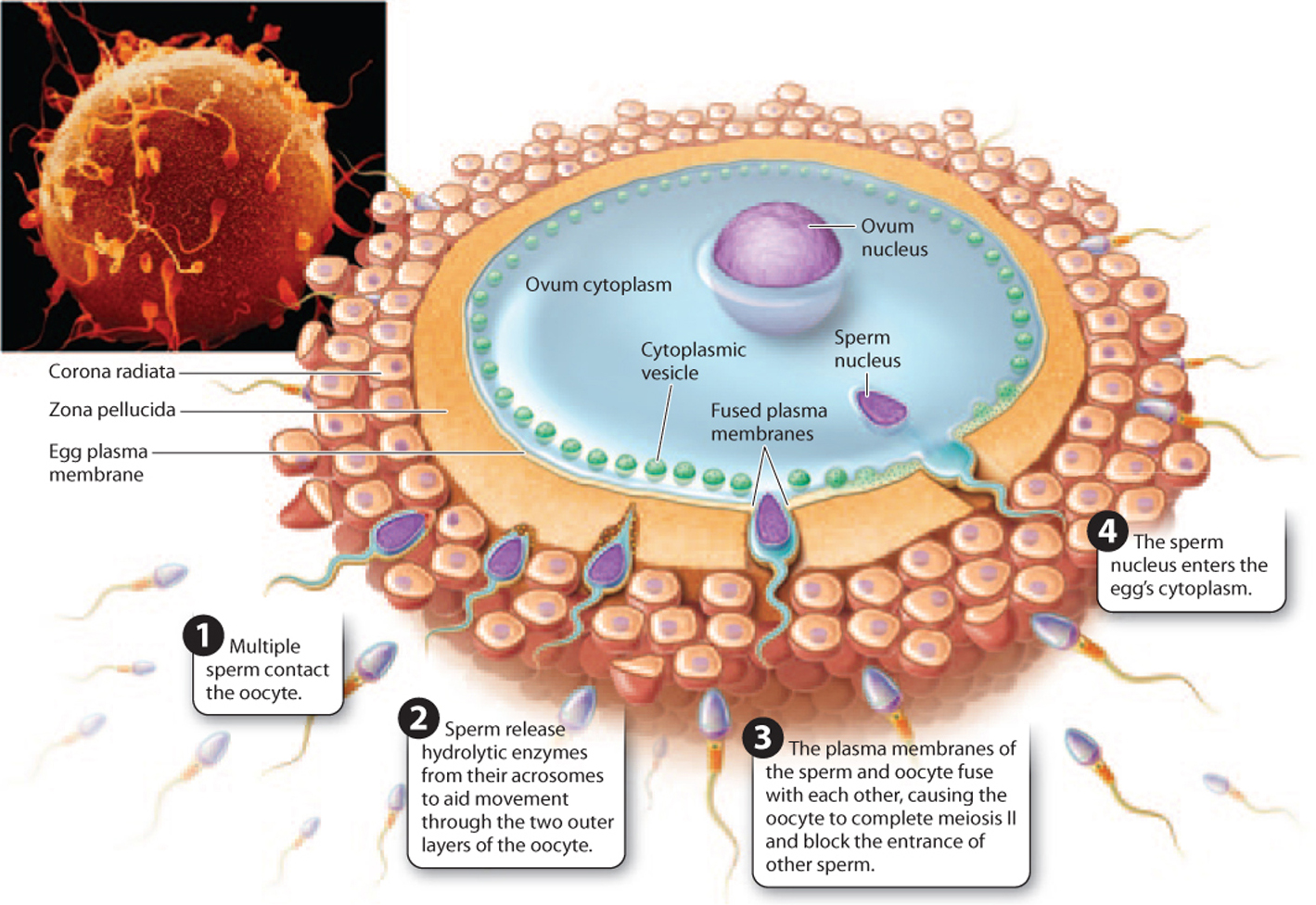
In many species, the oocyte releases soluble molecules, creating a concentration gradient that sperm use as a guide to find the oocyte. On reaching the oocyte, the sperm passes through two layers—an outer layer of follicle cells called the corona radiata and an inner matrix of glycoproteins called the zona pellucida—before fusing with the plasma membrane of the oocyte (Fig. 42.20). In sea urchins, by contrast, there is a single vitelline membrane surrounding the egg. To facilitate its passage, the head of the sperm releases enzymes from the acrosome. In addition, surface proteins on the gametes allow them to interact in a species-specific manner. For organisms with external fertilization, such as sea urchins, these proteins are especially important.
After passing through the layers surrounding the ooctye, the plasma membranes of the sperm and oocyte fuse (Fig. 42.20). The oocyte then completes meiosis II and changes occur that prevent polyspermy, or fertilization by more than one sperm. These mechanisms are classified according to how quickly they act and have been extensively studied in sea urchins. The first line of defense, or fast block to polyspermy, is caused by changes in the membrane potential of the egg, which is usually negative but quickly becomes positive on fertilization. The fast block is quick but temporary and does not occur in mammals.
There is also a second, more stable defense, or slow block to polyspermy. In this process, vesicles in the oocyte fuse with the plasma membrane and release their contents, leading to modification of the structure of the zona pellucida in mammals and the vitelline membrane in sea urchins. The fertilized, diploid egg is a zygote—the first cell in the development of a new organism.
Although sexual intercourse is one way that gametes are brought together, there are now a variety of other methods to achieve fertilization and pregnancy in humans. These are collectively called assisted reproductive technologies and are typically used to treat infertility. For example, in vitro fertilization (IVF) is a process in which eggs and sperm are brought together in a petri dish, where fertilization and cell divisions occur. The developing embryos are then inserted into the uterus. The first IVF or “test-tube” baby was born in 1978, and since then several million babies have been conceived in this way. Assisted reproductive technologies such as IVF have opened the door to pre-implantation genetic diagnosis, in which oocytes or embryos are screened for genetic diseases before implantation. This technique raises ethical concerns, as it can be used to determine and select the gender of the embryo and other traits not associated with disease.
42.4.3 The first trimester includes cleavage, gastrulation, and organogenesis.
How does a single cell become a complex, multicellular organism? Development depends on a precisely orchestrated set of cell divisions, movements, and specializations coordinated by both maternal and zygotic genetic instructions (Chapter 20). In this section, we focus on three key processes in early embryonic development: cleavage, gastrulation, and organogenesis.
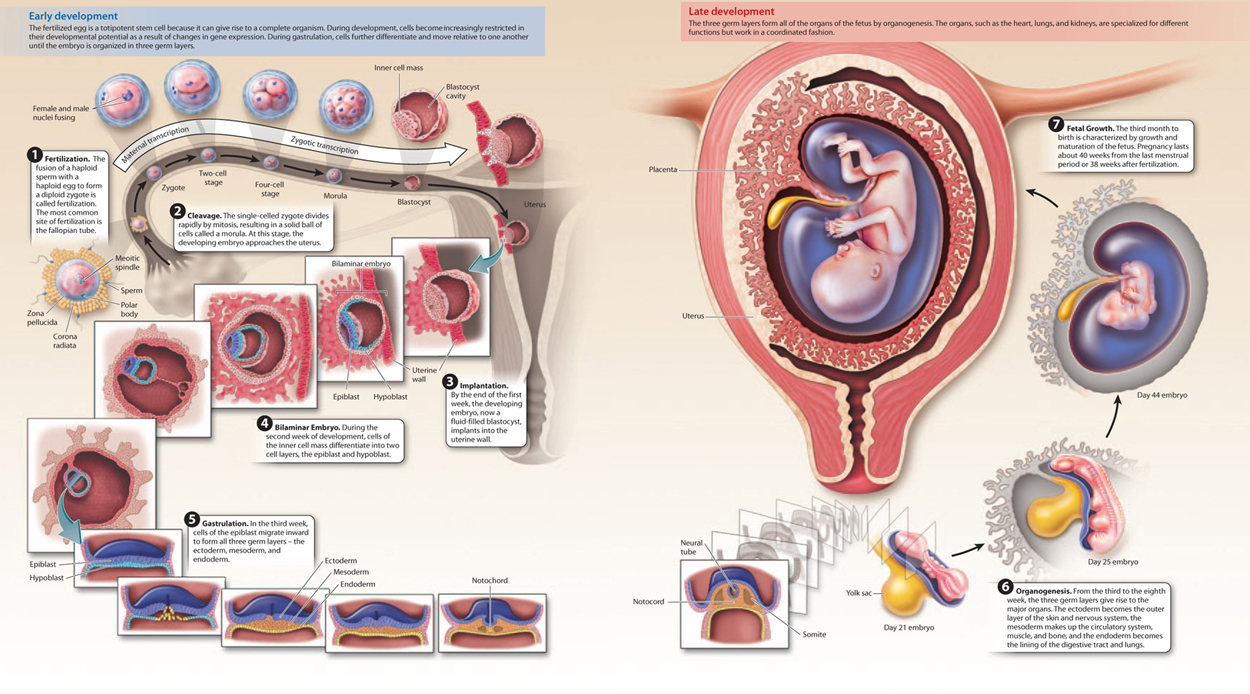
After fertilization, the single-celled zygote divides by mitosis, a process called cleavage (Fig. 42.21). This process divides the single large egg into many smaller cells. In most organisms, these mitotic divisions are rapid, occur at the same time, and are not accompanied by an overall increase in size of the embryo. Furthermore, they occur in the absence of expression of the zygote’s genes. Instead, molecules in the egg’s cytoplasm, including mRNA and protein encoded by maternal effect genes (Chapter 20) and stored in the egg, direct these early cell divisions.
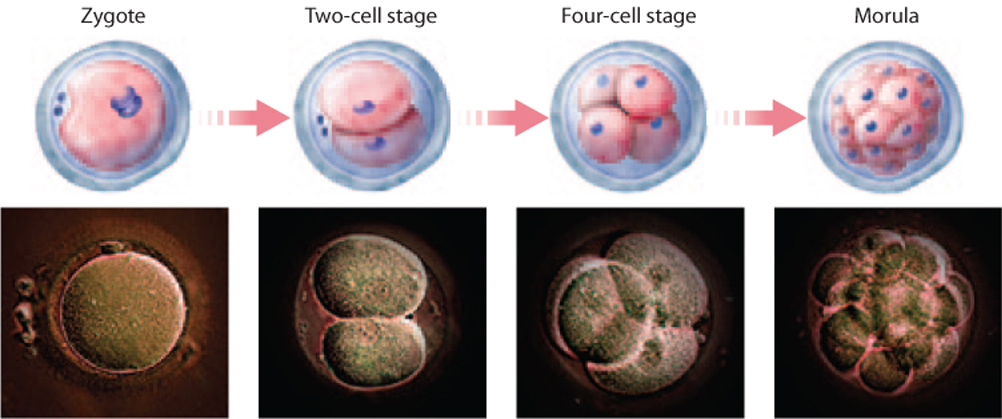
Cleavage in mammals is unique in several respects. It is slow compared to cleavage in other organisms. In addition, in mammals the early divisions do not occur at the same time, so it is not unusual to find an odd number of cells in the developing embryo. Finally, zygotic transcription begins very early, as early as the first mitotic division, or two-cell stage.
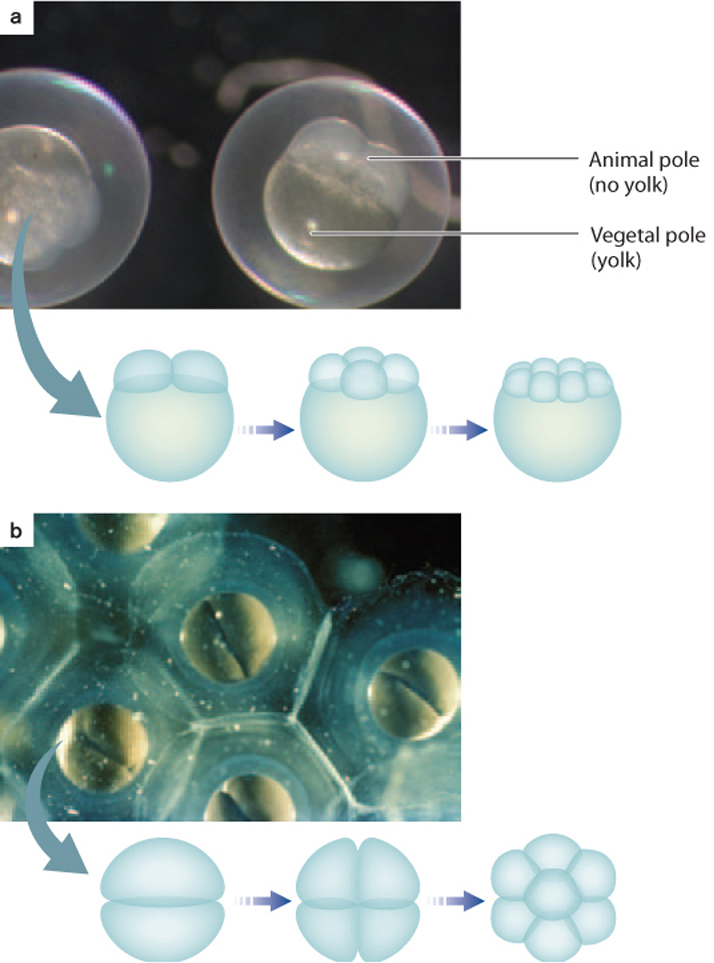
The pattern of cleavage is different for different organisms and is determined in part by the amount of yolk present in the egg (Fig. 42.22). In animals with lots of yolk, such as insects, fish, and birds, the embryo has two poles—a vegetal pole, where the yolk is concentrated, and an animal pole, where it is less concentrated. The presence of yolk inhibits cell division. Therefore, in these animals, cell division does not extend through the entire egg (Fig. 42.22a). In animals with very little yolk, such as mammals and sea urchins, cell division extends completely through the egg (Fig. 42.22b). In the absence of yolk, the developing embryo needs another source of nutrition, which in humans and other placental mammals is provided through the placenta.
Cleavage results in a solid ball of cells called a morula (see Fig. 42.21). Further cell divisions result in a fluid-filled ball of cells called a blastula. It is around this time, about 5 days after fertilization, that the developing embryo implants into the uterine lining, which is thickened and highly vascularized under the influence of progesterone from the corpus luteum.
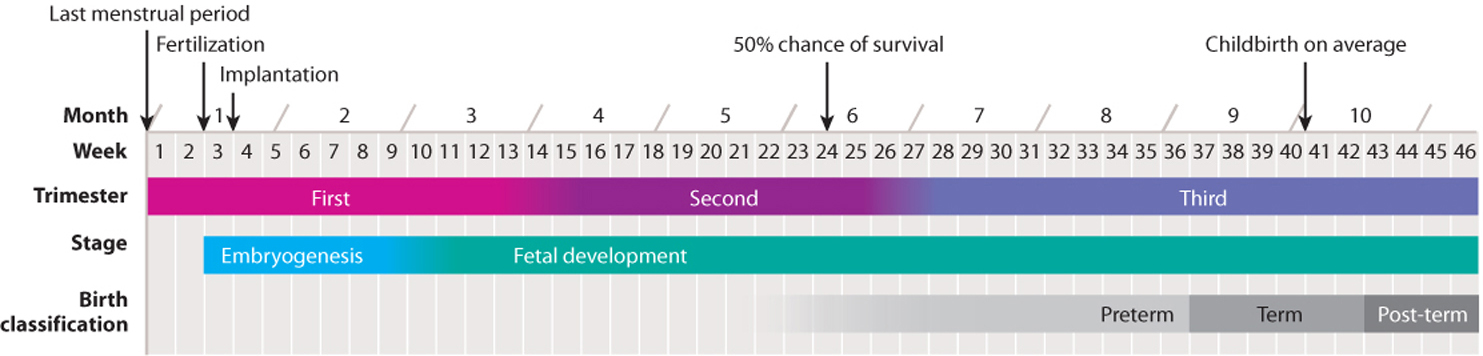
Implantation marks the beginning of pregnancy, the carrying of one or more embryos in the uterus. It lasts approximately 38 weeks from fertilization. Because the time of fertilization is often not known, human pregnancy is typically measured as 40 weeks from the last menstrual period. Pregnancy is divided into three periods called trimesters, each lasting about 3 months (Fig. 42.23). The divisions between the three trimesters are arbitrary, but they provide a convenient way to follow the major changes that occur during pregnancy.
The first trimester is characterized by spectacular changes in the developing embryo under the influence of genes and developmental pathways that are largely conserved across vertebrates (Chapter 20). During this time, the embryo develops all its major organs and begins to be recognizably human. Toward the end of the first trimester, a human embryo is known as a fetus.
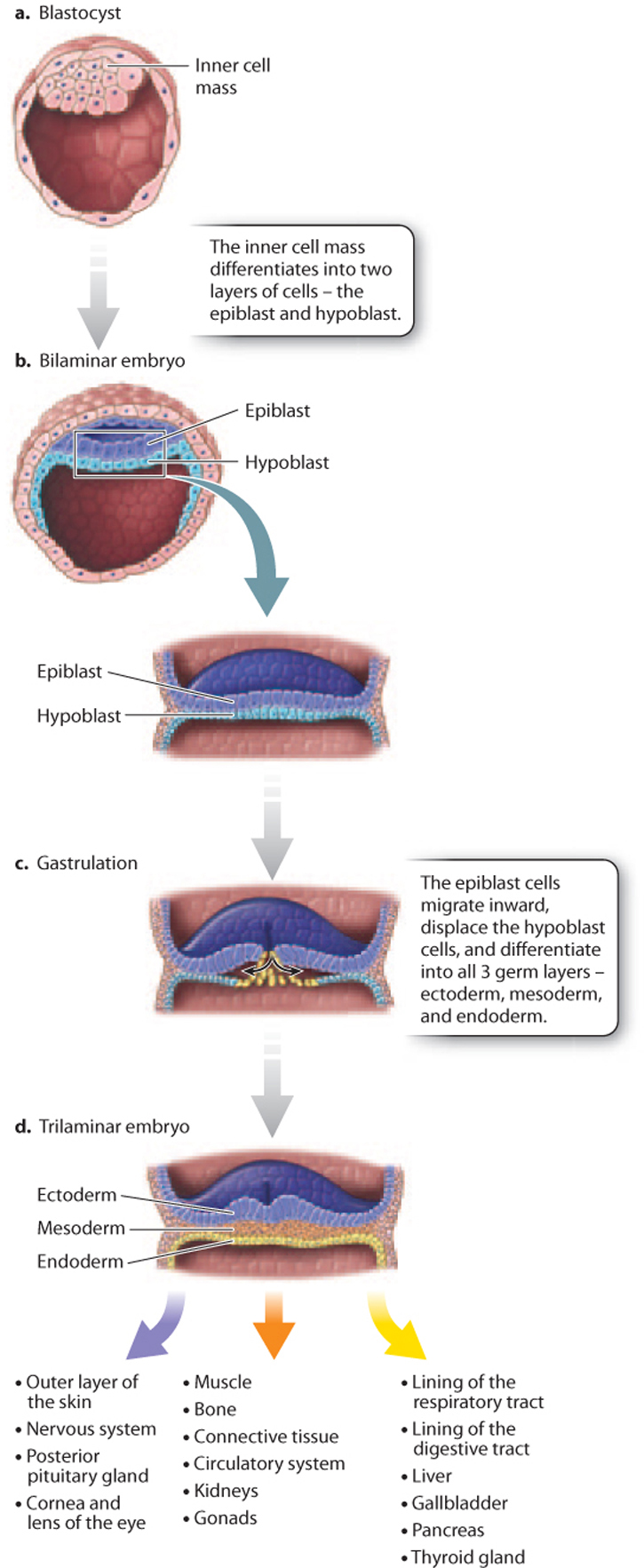
In mammals, such as humans, the blastula has an inner cell mass and an outer layer of cells, and is known as a blastocyst (Fig. 42.24a). The inner cell mass becomes the embryo itself, and the outer cell layer forms part of the placenta. The placenta is composed of both maternal and embryonic cells and is the site of gas exchange, nutrient uptake, and excretion of waste between maternal and fetal blood. The placenta also secretes the hormones estrogen and progesterone, which maintain the uterine lining during pregnancy. Meanwhile, the inner cell mass differentiates into a two-layered (bilaminar) structure consisting of the epiblast and hypoblast (Fig. 42.24b).
Gastrulation is the second key developmental process in embryogenesis. It is a highly coordinated set of cell movements that leads to a fundamental reorganization of the embryo. In humans, cells of the epiblast divide and migrate inward (Fig. 42.24c). These migrating epiblast cells form three layers or sheets of cells called germ layers, which include the outer ectoderm, intermediate mesoderm, and inner endoderm (Fig. 42.24d). All of the adult tissues and organs are made from the three germ layers. At this stage, the embryo is known as a trilaminar (three-layered) embryo to reflect the three germ layers.
As with cleavage, the details of gastrulation vary in different organisms. One notable difference is the fate of the opening through which cells migrate during gastrulation. In protostomes (“mouth first”), such as worms, mollusks, and arthropods, it becomes the mouth, whereas in deuterostomes (“mouth second”), such as vertebrates and sea urchins, it becomes the anus and the mouth forms later (Chapter 44).
Organogenesis, the third key developmental process, is the transformation of the three germ layers into all the organ systems of the body. The ectoderm becomes the outer layer of the skin and nervous system, the mesoderm makes up the circulatory system, muscle, and bone, and the endoderm becomes the lining of the digestive tract and lungs (Fig. 42.24d). Because all the basic structures and organ systems develop during the first trimester of pregnancy, it is during this time that the developing fetus is most vulnerable to toxins, drugs, and infections.
It is also during the first trimester that the embryo begins to develop male or female characteristics. In humans, sex determination is achieved by the presence or absence of the Y chromosome. The Y chromosome contains a gene, SRY, that encodes a protein called testis-determining factor (TDF). TDF in turn acts on the developing gonad to cause it to differentiate into a testis. In the absence of TDF (in females), the immature gonad becomes an ovary. Once sex is determined, sexual differentiation follows under the influence of hormones produced by the gonads. In humans, then, sex determination is genetic, as is the case in other mammals and in birds. For other organisms, such as some reptiles and fish, sex determination depends instead on environmental factors, relying on such cues as temperature and crowding.
42.4.4 The second and third trimesters are characterized by fetal growth.
The second trimester, from 13 to 27 weeks, is characterized by continued development and growth of the fetus. This is the time when fetal movements can first be felt by the mother. Babies born in the last few weeks of the second trimester are often able to survive, and are considered premature. Because of improvements in the intensive care of premature babies, babies born as early as 22 weeks after fertilization can survive.
The third trimester, from 28 to 40 weeks, is characterized by rapid growth, with the fetus doubling in size during the last two months. Weight gain is critical, as birth weight is a major factor in infant mortality. The fetus often moves less frequently during this trimester as it becomes increasingly compressed by limited space. Although 40 weeks (from the last menstrual period) is the average length of pregnancy, a baby born between 37 and 42 weeks is considered term. Human development from fertilization to the end of pregnancy is summarized in Fig. 42.25.
Quick Check 5
A multicellular organism starts off as a single cell, which divides by mitosis to produce many cells. Therefore, for the most part, all the cells in a multicellular organism are genetically identical—they are clones—yet in time different groups of cells look different and function quite differently from one another. How can this be?
Visual Synthesis: Reproduction and Development
Click the image below to view an enlarged version in a new window.
42.4.5 Childbirth is initiated by hormonal changes.
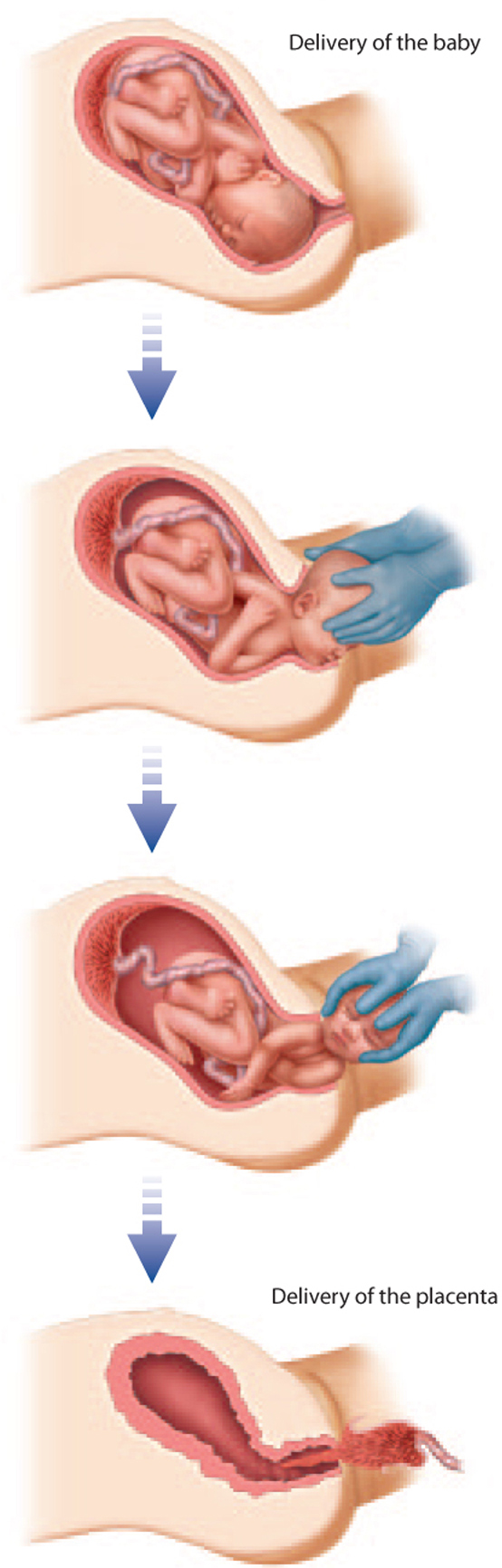
Childbirth marks the end of pregnancy. Although we tend to think of childbirth as the delivery of the baby, it actually consists of three events—changes in the cervix to allow passage of the baby, descent and delivery of the baby, and delivery of the placenta. These three events result from strong muscular contractions of the uterus mediated by the hormone oxytocin from the posterior pituitary gland. Oxytocin causes uterine contractions, which stimulate more oxytocin release, which in turn leads to further contractions, and so on.
During pregnancy, the cervix is a tube about 4 cm long closed with a mucus plug. To allow passage of the infant, the cervix shortens and widens. These changes are not enough to accommodate passage of the baby during delivery. The head and shoulders of the fetus are wide relative to the size of the pelvis. Humans have a narrow pelvis relative to that of other primates as an adaptation for bipedalism. How, then, is the baby born? The baby goes through a series of movements that facilitate its passage through the pelvis (Fig. 42.26). Babies are usually born head first with the face pointing down (toward the mother’s back) when passing through the birth canal. Other positions make vaginal birth more difficult, sometimes impossible.
The third and final stage of childbirth is the delivery of the placenta, which usually occurs about 15 to 30 minutes after the baby is born.
Although birth normally occurs vaginally, in some cases, such as when the baby is too large or positioned feet first, it is necessary to remove the baby by making an incision in the abdomen and uterus in a surgical procedure known as a Caesarian section. Interventions such as Caesarian sections, the use of antibiotics, and pre- and postnatal care have greatly reduced maternal and fetal mortality associated with childbirth.
Whichever way the baby is delivered, when the umbilical cord is cut and the baby takes its first breath, it begins its life independent of the mother—anatomically independent, that is. Otherwise, human newborns are completely dependent on others. Humans are unusual among animals in having an extended period of helplessness and immaturity while their brains and bodies grow and develop. This time means that parents usually put considerable effort into raising and caring for their offspring, and it allows for extended periods of learning, the development of strong social bonds, and the transmission of culture to the next generation.