44.5 THE EVOLUTIONARY HISTORY OF ANIMALS
Because many animals form mineralized skeletons, sedimentary rocks deposited during the past 542 million years contain a rich fossil record of shells and bones. Animals also leave a sedimentary calling card in their tracks, trails, and burrows, again widespread in sedimentary rocks formed after 542 million years ago. Not all animal phyla are well represented in the fossil record—fossil annelids, for example, are rare and flatworms are essentially unknown as fossils because they lack mineralized body parts (Chapter 23). In contrast, the fossilized shells and bones of bivalve mollusks, brachiopods, echinoderms, and mammals preserve an excellent record of evolutionary history within these groups. If we step back and look at the big picture, what do fossils tell us about the evolutionary history of the animal kingdom?
44.5.1 Fossils and phylogeny show that animal forms were initially simple but rapidly evolved complexity.
Phylogeny suggests that animals are relative latecomers in evolutionary history, and the fossil record confirms this hypothesis. Life originated more than 3.5 billion years ago, but, as discussed in previous chapters, microorganisms dominated ecosystems for most of our planet’s history. As noted in Chapter 28, macroscopic fossils of organisms thought to be animals first appear in rocks deposited only 575 million years ago. Called Ediacaran fossils after the Ediacara Hills of South Australia where they were discovered, these fossils have simple shapes that are not easily classified among living animal groups (Fig. 44.37; see also Fig. 28.13).

Phylogeny suggests that we should look for sponges, cnidarians, and other diploblastic animals among the oldest animal fossils, but sponges, at least, are rare among Ediacaran fossils. Instead, a majority of Ediacaran fossils show simple, fluid-filled tubes, without identifiable mouths or other organs. Many may have formed colonies, gaining complexity through colonial growth and differentiation, as some living cnidarians do. These early animals had epithelia and are thought to have obtained food by taking in dissolved organic matter while exchanging gases by diffusion, somewhat like living placozoans. Most Ediacaran fossils probably form an early branch or branches on the animal tree, falling among the limbs whose survivors include cnidarians, ctenophores, and placozoans (Fig. 44.37).
Why did the first animal radiation occur so late in the evolutionary day? Scientists continue to debate this question, but part of the answer appears to lie in Earth’s environmental history. Geochemical data suggest that only during the Ediacaran Period did the atmosphere and oceans come to contain oxygen in quantities sufficient to support the metabolism of large, active animals.
44.5.2 The animal body plans we see today emerged during the Cambrian Period.
Ediacaran fossils differ markedly from the shapes of living animals, but in the next interval of geologic history, the Cambrian Period (542–489 million years ago), we begin to see the remains of animals with familiar body plans (Fig. 44.38). Cambrian fossils commonly include skeletons made of silica, calcium carbonate, and calcium phosphate minerals, and these record the presence of arthropods, echinoderms, mollusks, brachiopods, and other bilaterian animals in the oceans. Spicules made by sponges are common in Cambrian rocks as well. The rocks also preserve complex tracks and burrows made by organisms with hydrostatic skeletons, muscular feet, and the jointed legs of arthropods. And in a few places, notably at Chengjiang in China and the Burgess Shale in Canada, unusual environmental conditions have preserved a treasure trove of animals that did not form mineralized skeletons (Chapter 23).

These exceptional windows on early animal evolution show that, during the first 40 million years of the Cambrian Period, the body plans characteristic of most bilaterian phyla took shape in a transition sometimes called the Cambrian explosion. Sponges and cnidarians radiated as well, producing through time the biomass- and diversity-enhancing habitats of reefs and imparting an ecological structure to life in the sea broadly similar to what we see today.
Scientists sometimes argue about whether the name “Cambrian explosion” is apt. The fossil record makes it clear that bilaterian body plans did not suddenly appear fully formed, so the event was not truly “explosive.” Rather, fossils demonstrate a huge accumulation of new characters in a relatively short period of time, during which the key attributes of modern animal phyla emerged. For example, living arthropods combine segmented bodies with a protective cuticle, jointed legs, other appendages specialized for feeding or sensing the environment, and compound eyes with many lenses. Cambrian fossils include the remains of organisms with some but not all of the major features combined today in arthropods. In short, the first 40 million years of Cambrian animal evolution ushered in a world utterly distinct from anything known in the preceding 3 billion years.
Despite the burst of evolution recorded by Cambrian fossils, there were still relatively few species of marine animals at the end of the Cambrian Period. The ensuing Ordovician Period (489–444 million years ago) was a time of renewed animal diversification, especially the evolution of heavily skeletonized animals in the world’s oceans. The Ordovician radiation established a marine ecosystem that persisted for more than 200 million years. Interestingly, however, if you had walked along an Ordovician beach, the shells washing about your feet would have been far different from the ones you see today. The dominant shells were those of brachiopods, not clams. Broken corals in the surf were the skeletons of now-extinct cnidarians only distantly related to modern reef-forming corals. And arthropod shells, molted during growth, were those of now-extinct trilobites, not lobsters or crabs. This ecosystem was devastated by a mass extinction 252 million years ago (Chapter 23).
44.5.3 Animals began to colonize the land 420 million years ago.
The colonization of land by animals began only after plants had established themselves there. Land plants evolved about 460 million years ago. Arthropods followed by about 420 million years ago, especially chelicerates that included the ancestors of spiders, mites, and scorpions. Insects, descended from crustacean ancestors that independently gained access to the land surface, appeared at about the same time. The radiation of the major groups of insects, however, began 360 million years ago with a marked diversification of dragonflies and the ancestors of cockroaches and grasshoppers. Some of the dragonfly-like insects that darted among the plants of Carboniferous coal swamps had bodies the size of a lobster and 75-cm wing spans! One might well wonder what prevents insects from attaining this size today. These immense insects flew through an atmosphere that contained more oxygen than is present today, and this higher level of atmospheric oxygen is thought to have been a necessary condition for insect gigantism. After some time, levels of atmospheric oxygen declined to levels closer to those seen today, and the giant insects disappeared. Flies, beetles, bees, wasps, butterflies, and moths radiated later, beginning in the early Mesozoic Era. Their rise in diversity parallels that of the flowering plants and reflects coevolution of these pollinators and flowering plants.
Tetrapod fossils first appear in sedimentary rocks deposited near the end of the Devonian Period, about the same time as the early insect radiations 360 million years ago. As discussed in Chapter 23, the fossil record documents in some detail the shifts in skull, trunk, and limb morphology that allowed vertebrate animals to colonize land. These include the evolution of muscled, articulated legs from fins, together with a set of strong, articulated digits where the limbs meet the ground, internal lungs rather than gills, and an erect, elevated head with eyes oriented for forward vision. The appearance of amphibians and then reptiles follows the predictions of phylogenies based on comparative biology.
The most remarkable animals among the reptiles are surely the dinosaurs. Dinosaurs first evolved about 210 million years ago from small, bipedal ancestors that radiated following the mass extinction 252 million years ago. Once established, the dinosaurs radiated to produce many hundreds of species, dominating terrestrial ecosystems until 65 million years ago, when nearly all dinosaur species disappeared, the most familiar victims of an enormous and Earth-changing asteroid impact (see Fig. 1.3). We say “nearly all” because the fossil record documents the divergence of the ancestors of modern birds from a specific subgroup of dinosaurs, about 150 million years ago (see Fig. 23.20).
Mammals have been the dominant vertebrates in most terrestrial ecosystems since the extinction of dinosaurs, but the group originated much earlier, at least 210 million years ago. During the age of dinosaurs, most mammals were small nocturnal or tree-dwelling animals that stayed out of the way of large dinosaurs, although fossils from China show clearly that the largest mammals of this interval ate small dinosaurs and their eggs.
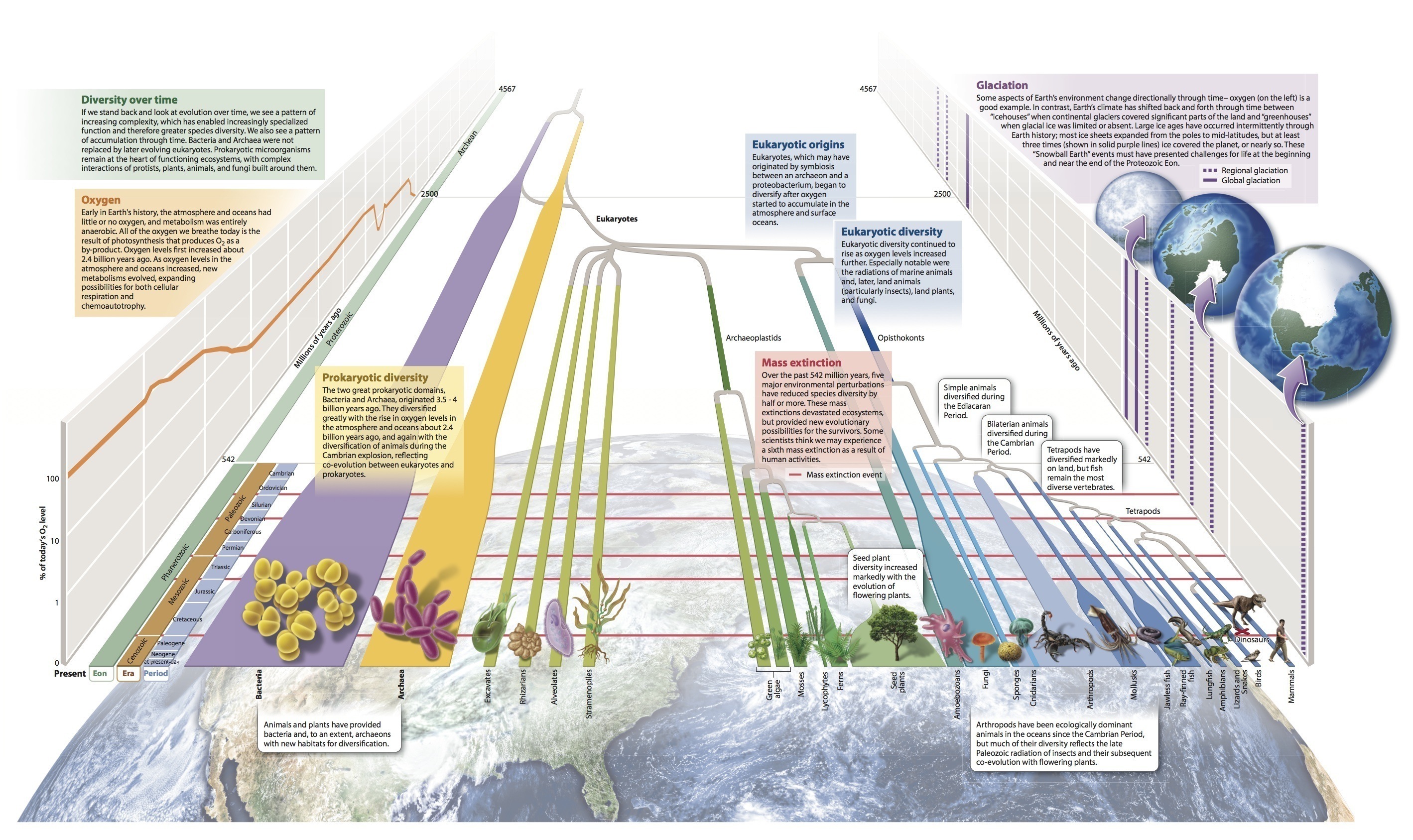
We summarize the biological diversity of all organisms, including animals, along with a timeline of Earth’s history, in Fig. 44.39.
Case 8 Biodiversity Hotspots: Rain Forests and Coral Reefs
44.5.4 How have coral reefs changed through time?
On land, tropical rain forests are hotspots of diversity, and coral reefs support unusually large numbers of species in the oceans. Reefs have existed through most of Earth’s recorded history, and rain forests have existed since the origin of trees more than 370 million years ago. Through time these have been Earth’s most diverse communities in the sea and on land. But the taxonomic composition of these communities has changed through time because of both evolutionary innovation and mass extinction.
Bacteria built the first reefs, layered mounds of limestone that rose above the seafloor. With the Cambrian diversification of animals, calcified sponges rose to ecological prominence as participants in reef construction, but these communities were short lived, as mass extinction about 515 million years ago essentially wiped out the reef-building sponges. Subsequent renewed diversification of sponges, bryozoans, and corals with massive calcium carbonate skeletons resulted in new and different reef communities beginning in the Ordovician Period. Eventually these communities were also destroyed by mass extinction during the Devonian Period. Reefs were widespread again by the Permian Period (300–252 million years ago), dominated this time by different groups of sponges and bryozoans, along with skeleton-forming green algae.
The devastating extinction at the end of the Permian Period removed all corals and calcifying sponges, and for several million years following this event, reefs were built only by microorganisms, much as they were before the evolution of animals. Then, the evolution of new skeleton-forming corals (from sea anemone-like ancestors) and a resurgence of sponges capable of massive calcification introduced new reef systems into the ocean. Given the iconic status of coral reefs as examples of marine biodiversity, it is important to note that reefs like those of the present day have developed only over 40 million years of Earth history.
As discussed in Chapter 48, we may be nearing another great change in reef biology as global climate change threatens reef communities in many parts of the world. Today’s coral species are at their tolerance limits of ocean temperature and pH, and both may be changing more rapidly than species can adapt. From the perspective of Earth history, the loss of coral reefs would simply be one more in a long series of extinction episodes. For humans, however, the commercial and ecological consequences would be substantial and permanent.
Visual Synthesis: Diversity through Time
Click the image below to view an enlarged version in a new window.