Chapter 1. Population Growth
Introduction
In order to understand the concepts of population growth quantitatively, we need some sort of mathematical notation. For the sake of simplicity, we will minimize the number of symbols while carefully defining each as we move along. Note however, that biologists in different specialties often use different symbols for the same concepts. Hence if you consult a textbook in ecology, population biology, demography, or epidemiology, or look at an internet site in one of these fields, you may find a different mathematical notation than that used here.
Discrete Population Growth
To obtain a mathematical expression for population growth, we must first formulate a model or idealized population of organisms that specifies exactly when during the life cycle the organisms reproduce. The simplest population-growth model is that of an organism with a life cycle analogous to that of an annual plant, in which each generation of organisms germinates at the same time, grows together to sexual maturity, reproduces simultaneously, and then dies. As time goes on, the number of germinating seeds produced by organisms in any one generation determines the number of organisms in the succeeding generation. This model of population growth is known as discrete population growth because the population size increases (or decreases) in one discrete step at the beginning of each new generation. In principle, we should also take immigrants into the population into account, as well as emigrants who leave the population, but here we will assume that the population is isolated with no immigration or emigration.
Mathematically, suppose we let Nt represent the number of organisms present in the population in generation t, where t can take on any integer value 0, 1, 2,.... (N0 therefore represents the number of organisms present in the original, or 0th generation.) We can then formulate population growth by the equation Nt = Nt-1(1 + r), which when put into words says that the number of organisms present in any generation, Nt, equals the number present in the previous generation (Nt-1) times the average number of offspring (1 + r) that each of these Nt-1 parents produce. In this equation, r represents the rate of population growth. When r is negative, the population decreases in size, when r is positive it increases in size, and when r is 0 it doesn’t change. In the real world, r can change from one generation to the next, but for starters we will assume that r is constant.
Note from the equation that r can also be written as r = (Nt – Nt-1)/ Nt-1, which says that, in the discrete model of population growth, r is the proportional growth (or decline) in population size in any one generation. We emphasize that the quantity r is a rate – a change per unit of time. In the discrete model, the unit of time is "per generation." In other words, in the discrete model, r is the proportionate change in population size [(Nt – Nt-1)/ Nt-1] per generation. Some biologists like to use the following definition: In the discrete model of population growth, r is the percentage increase (or decrease) in population size, expressed in decimal form, per generation.
The equation Nt = Nt-1(1 + r) implies that N1 = N0(1 + r). However, since the relation between N2 and N1 is the same as that between N1 and N0, we can write N2 = N1(1 + r) = N0(1 + r)((1 + r) = N0(1 + r)2. Likewise N3 = N2(1 + r) = N0(1 + r)3, and continuing in this manner, we obtain the general solution Nt = N0(1 + r)t.
Continuous Population Growth
In an alternative model to discrete population growth, the organisms' life cycles are not synchronized in time. Organisms are born and develop at different times, and any organism that reaches sexual maturity in the population can reproduce immediately. The result is that the population does not increase or decrease in discrete steps, but continuously. This type of population growth is known as continuous population growth, and a mathematical formulation for population size N at any time t, symbolized N(t), is given by dN(t)/dt = rN(t). Here we use N(t) to emphasize that t can take on any nonnegative value, not merely integer values. The expression dN(t)/dt is the instantaneous rate of change of N(t) at time t. (For those familiar with calculus, dN(t)/dt is the derivative of the function N(t) at time t.) In the continuous model of population growth, r is known as the intrinsic rate of increase (or decrease) in population size. The solution to the equation for continuous growth is N(t) = N(0)ert, where N(0) represents the initial population size (at time t = 0) and e is the base of natural logarithms and equals approximately 2.718. As in the discrete model, the population is shrinking, staying the same, or growing in size according to whether r < 0, r = 0, or r > 0.
What is the relation between the discrete and continuous models? It turns out that, when r is sufficiently small, the population size in the continuous model N(t) = N(0)ert, for integer values of t, is approximately the same as that in the discrete model Nt = N0(1 + r)t. The reason for the agreement is a mathematical relation stating that er = 1 + r + r2/2! + r3/3! + ..., where the symbol ! stands for factorial (the product of all integers up to and including the number preceding the factorial sign). Although in principle the sum goes on for infinitely many terms, when r is small enough that r2 is much less than r (in practical terms, this means r = 0.01 or smaller), then er = 1 + r approximately. But since ert = (er)t, then ert = (1 + r)t approximately. In this case, N(t) = N(0)ert can be written as approximately N(t) = N(0)(1 + r)t, and this equation is analogous to Nt = N0(1 + r)t that we derived earlier in the discrete model of population growth.
The accompanying graph compares discrete and continuous population growth in the case r = 0.01 starting from an initial population size of 10 individuals and extending over 100 generations. The red curve is the exact solution in the continuous model [N(t) = N(0)ert] and the blue dots are the exact values in the discrete model [Nt = N0(1 + r)t].

The good fit over this many generations reflects the fact that, when r = 0.01, the approximation ert = (1 + r)t is very good. Bear in mind, though, that if r is too large, or if the time scale is too long, the approximation ert = (1 + r)t breaks down.
What Units For Time?
In the continuous model, r is the instantaneous rate of increase in N(t) at time t. (Technically, it is the slope of a straight line tangent to the curve N(t) at time t.) This definition is problematic because one cannot measure population growth at an "instant" in time, and the definition is also not of much use to a hospital administrator wanting to know how many babies are likely to be born next year or to a wildlife resource specialist wanting to know the size of next year’s deer population. On the other hand, suppose you could choose an interval of time that is small relative to the generation time of the organism (for bacteria this might be per minute, or for other organisms per day), and could estimate the population size at the beginning and end of such an interval. To be concrete, let us symbolize the change in population size over the time interval as ΔN and the length of the time interval as Δt. Then analogous to dN(t)/dt = rN(t) we could write
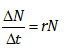
where N is the population size at the beginning of the interval. Then, dividing through by N, we can define
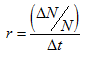
This definition of r is different from that defined in both the discrete model (where r was per generation) or the continuous model (where r was instantaneous). The r based on ΔN and Δt is the average rate of population growth per unit time over the interval Δt. But if the time interval Δt is small relative to the generation time of the organism, then the average rate of population growth over Δt is approximately equal to the instantaneous (intrinsic) rate of increase in the continuous model. For example, an estimate of the maximum intrinsic rate of increase in the human population is approximately 0.0003 per day. (This is the rate at which the human population could increase in size under ideal conditions with everyone reproducing as rapidly as possible.) With r = 0.0003/day, the human population size would double every 6.3 years! Mercifully, most actual human population growth rates are only 10% or less of the maximum.
Logistic Growth
The example of how rapidly the human population could grow if it were increasing at its maximum rate is instructive because it demonstrates that no population can grow in size at its maximum rate for long periods of time, otherwise it would consume all available resources or fill all available space. As another example of unrestricted growth, consider that a single cell of the bacterium Escherichia coli, under ideal conditions, can undergo division in about 20 minutes. If a single cell continued to divide at this rate, with the population of bacterial progeny steadily increasing, then after only 30 hours the volume of the bacterial population would equal the volume of all the world's oceans combined!
Realistic models of population growth must therefore also include some upper limit defined by the availability of resources or space. Again we must start by defining a model specifying exactly how population growth is limited, and again we can consider organisms that reproduce at discrete intervals (discrete model) or those that reproduce continuously in time (continuous model). A simple but nevertheless useful model for incorporating a growth limit is to assume that the growth rate decreases gradually as population size increases until some population size is reached at which the growth rate becomes 0. In other words, instead of using r as a measure of growth rate, as in the models discussed earlier, we use r(K – N)/K, where N is the current population size. When N is much smaller than K, then r(K – N)/K is very close to r; but as N increases and becomes closer and closer to K, then r(K – N)/K gradually decreases; and when N = K the population growth rate becomes 0. The constant K is usually called the carrying capacity of the environment because it determines the upper limit of population size. Population growth limited in this manner is call logistic growth. ("Logistic" is an old term having to do with mathematical calculation.)
In a discrete population growing at a rate of r(K – N)/K per generation, the logistic equation for population size Nt in generation t is given by
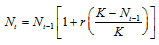
This equation is not so easily solved as the one we solved earlier for unlimited growth, but for any given values of N0, r, and K, it is relatively easy to write a computer program that will calculate N1, N2, N3, … for however many generations may be desired.
For a population growing continuously in time, the equation corresponding to logistic growth is
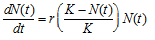
which does have a relatively simple solution, namely
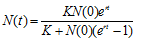
Logistic growth in a continuous model is shown in the graph below for the values r = 0.01, K = 100, and N(0) = 10.

The S-shaped, sigmoidal curve is characteristic of logistic population growth, which starts out at an exponential rate but then decreases gradually to 0 as the population size approaches the carrying capacity K.