3.2 Vision
FROM LIGHT TO SIGHT

KEY THEME
The receptor cells for vision respond to the physical energy of light waves and are located in the retina of the eye.
KEY QUESTIONS
What is the visible spectrum?
What are the key structures of the eye and their functions?
What are rods and cones, and how do their functions differ?
A lone caterpillar on the screen door, the pile of dirty laundry in the corner of the closet, a spectacular autumn sunset, the intricate play of color, light, and texture in a painting by Monet. The sense organ for vision is the eye, which contains receptor cells that are sensitive to the physical energy of light. But before we can talk about how the eye functions, we need to briefly discuss some characteristics of light as the visual stimulus.
What We See
THE NATURE OF LIGHT
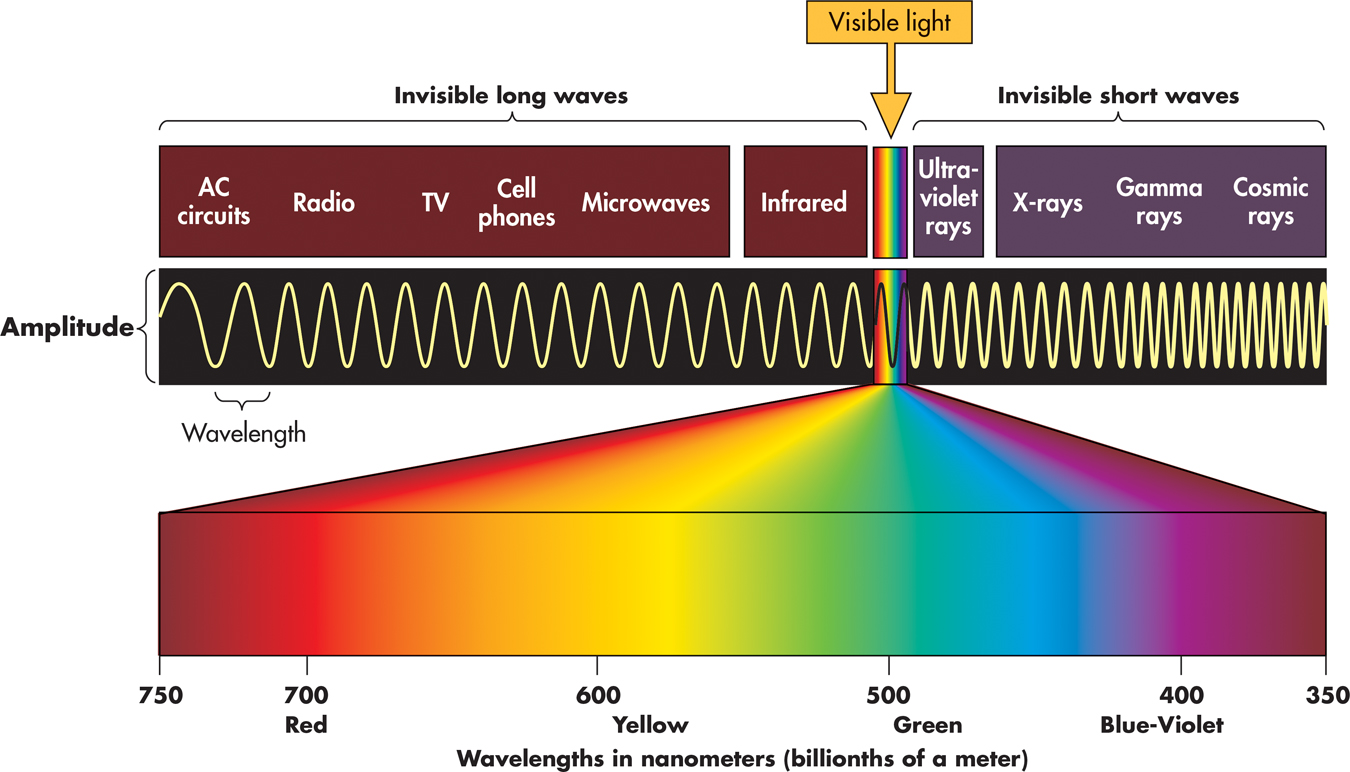
Light is just one of many different kinds of electromagnetic energy that travel in the form of waves. Other forms of electromagnetic energy include X-rays, the microwaves you use to pop popcorn, and the infrared signals or radio waves transmitted by your TV’s remote control. The various types of electromagnetic energy differ in wavelength, which is the distance from one wave peak to another.
wavelength
The distance from one wave peak to another.
Humans are capable of seeing only a minuscule portion of the electromagnetic energy range. In FIGURE 3.2, notice that the visible portion of the electromagnetic energy spectrum can be further divided into different wavelengths. As we’ll discuss in more detail later, the different wavelengths of visible light correspond to our psychological perception of different colors.
How We See
THE HUMAN VISUAL SYSTEM
Suppose you’re watching your neighbor’s yellow and white tabby cat sunning himself on the front steps. How do you see the cat? Simply seeing a yellow tabby cat involves a complex chain of events. We’ll describe the process of vision from the object to the brain. You can trace the path of light waves through the eye in FIGURE 3.3.
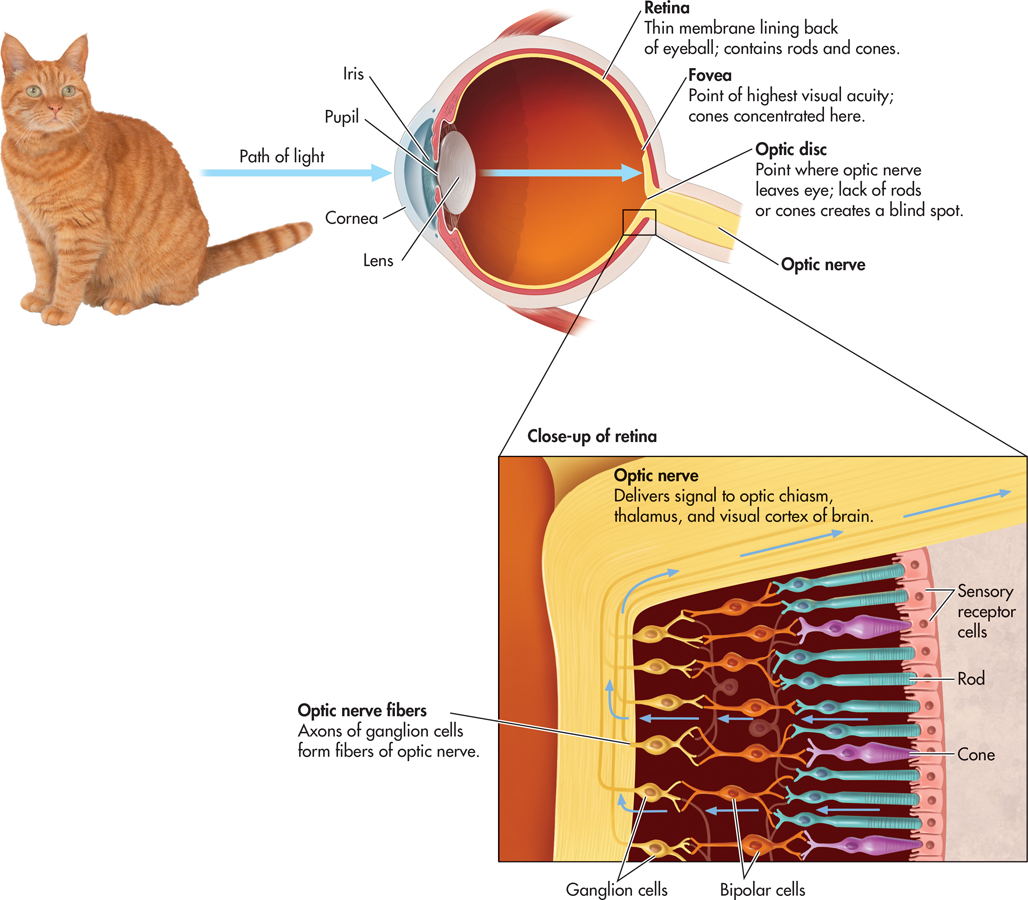
First, light waves reflected from the cat enter your eye, passing through the cornea, pupil, and lens. The cornea, a clear membrane that covers the front of the eye, helps gather and direct incoming light. The sclera, or white portion of the eye, is a tough, fibrous tissue that covers the eyeball except for the cornea. The pupil is the black opening in the eye’s center. The pupil is surrounded by the iris, the colored structure that we refer to when we say that someone has brown eyes. The iris is actually a ring of muscular tissue that contracts or expands to precisely control the size of the pupil and thus the amount of light entering the eye. In dim light, the iris widens the pupil to let light in; in bright light, the iris narrows the pupil.
cornea
(CORE-nee-uh) A clear membrane covering the visible part of the eye that helps gather and direct incoming light.
pupil
The opening in the middle of the iris that changes size to let in different amounts of light.
iris
(EYE-riss) The colored part of the eye, which is the muscle that controls the size of the pupil.
Behind the pupil is the lens, another transparent structure. In a process called accommodation, the lens thins or thickens to bend or focus the incoming light so that the light falls on the retina. If the eyeball is abnormally shaped, the lens may not properly focus the incoming light on the retina, resulting in a visual disorder. In nearsightedness, or myopia, distant objects appear blurry because the light reflected off the objects focuses in front of the retina. In farsightedness, or hyperopia, objects near the eyes appear blurry because light reflected off the objects is focused behind the retina. During middle age, another form of farsightedness often occurs, called presbyopia. Presbyopia is caused when the lens becomes brittle and inflexible. In astigmatism, an abnormally curved eyeball results in blurry vision for lines in a particular direction. Corrective glasses remedy these conditions by intercepting and bending the light so that the image falls properly on the retina. New surgical techniques, such as LASIK, correct visual disorders by reshaping the cornea so that light rays focus more directly on the retina.
lens
A transparent structure, located behind the pupil, that actively focuses, or bends, light as it enters the eye.
accommodation
The process by which the lens changes shape to focus incoming light so that it falls on the retina.
THE RETINARODS AND CONES
The retina is a thin, light-sensitive membrane that lies at the back of the eye, covering most of its inner surface (see FIGURE 3.3). Contained in the retina are the rods and cones. Because these sensory receptor cells respond to light, they are often called photoreceptors. When exposed to light, the rods and cones undergo a chemical reaction that results in a neural signal.
retina
(RET-in-uh) A thin, light-sensitive membrane, located at the back of the eye, that contains the sensory receptors for vision.
rods
The long, thin, blunt sensory receptors of the eye that are highly sensitive to light, but not to color, and that are primarily responsible for peripheral vision and night vision.
cones
The short, thick, pointed sensory receptors of the eye that detect color and are responsible for color vision and visual acuity.
Rods and cones differ in many ways. First, as their names imply, rods and cones are shaped differently. Rods are long and thin, with blunt ends. Cones are shorter and fatter, with one end that tapers to a point. The eye contains far more rods than cones. It is estimated that each eye contains about 7 million cones and about 125 million rods!
Rods and cones are also specialized for different visual functions. Although both are light receptors, rods are much more sensitive to light than are cones. Once the rods are fully adapted to the dark, they are about a thousand times better than cones at detecting weak visual stimuli (Masland, 2001). We therefore rely primarily on rods for our vision in dim light and at night.
Rods and cones also react differently to changes in the amount of light. Rods adapt relatively slowly, reaching maximum sensitivity to light in about 30 minutes. In contrast, cones adapt quickly to bright light, reaching maximum sensitivity in about 5 minutes. That’s why it takes several minutes for your eyes to adapt to the dim light of a darkened room but only a few moments to adapt to the brightness when you switch on the lights.
You may have noticed that it is difficult or impossible to distinguish colors in very dim light. This difficulty occurs because only the cones are sensitive to the different wavelengths that produce the sensation of color, and cones require much more light than rods do to function effectively. Cones are also specialized for seeing fine details and for vision in bright light.

Most of the cones are concentrated in the fovea, which is a region in the very center of the retina. Cones are scattered throughout the rest of the retina, but they become progressively less common toward the periphery of the retina. There are no rods in the fovea. Images that do not fall on the fovea tend to be perceived as blurry or indistinct. For example, focus your eyes on the word “For” at the beginning of this sentence. In contrast to the sharpness of the letters in “For,” the words to the left and right are somewhat blurry. The image of the outlying words is striking the peripheral areas of the retina, where rods are more prevalent and there are very few cones.
fovea
(FOE-vee-uh) A small area in the center of the retina, composed entirely of cones, where visual information is most sharply focused.
THE BLIND SPOT
One part of the retina lacks rods and cones altogether. This area, called the optic disk, is the point at which the fibers that make up the optic nerve leave the back of the eye and project to the brain. Because there are no photoreceptors in the optic disk, we have a tiny hole, or blind spot, in our field of vision. To experience the blind spot, try the demonstration in FIGURE 3.4.
optic disk
Area of the retina without rods or cones, where the optic nerve exits the back of the eye.
blind spot
The point at which the optic nerve leaves the eye, producing a small gap in the field of vision.
Why don’t we notice this hole in our visual field? The most compelling explanation is that the brain actually fills in the missing background information (Ramachandran, 1992a, 1992b; Weil & Rees, 2010). In effect, signals from neighboring neurons fill in the blind spot with the color and texture of the surrounding visual information (Supèr & Romeo, 2011).
!launch!

Processing Visual Information
KEY THEME
Signals from the rods and cones undergo preliminary processing in the retina before they are transmitted to the brain.
KEY QUESTIONS
What are the bipolar and ganglion cells, and how do their functions differ?
How is visual information transmitted from the retina to the brain?
What properties of light correspond to color perceptions, and how is color vision explained?
Visual information is processed primarily in the brain. However, before visual information is sent to the brain, it undergoes some preliminary processing in the retina by specialized neurons called ganglion cells. This preliminary processing of visual data in the cells of the retina is possible because the retina develops from a bit of brain tissue that “migrates” to the eye during fetal development (Hubel, 1995).
ganglion cells
In the retina, the specialized neurons that connect to the bipolar cells; the bundled axons of the ganglion cells form the optic nerve.
When the numbers of rods and cones are combined, there are over 130 million receptor cells in each retina. However, there are only about 1 million ganglion cells. How do just 1 million ganglion cells transmit messages from 130 million visual receptor cells?
VISUAL PROCESSING IN THE RETINA
Information from the sensory receptors, the rods and cones, is first collected by specialized neurons, called bipolar cells. Look back at the lower portion of FIGURE 3.3. The bipolar cells then funnel the collection of raw data to the ganglion cells. Each ganglion cell receives information from the photoreceptors that are located in its receptive field in a particular area of the retina. In this early stage of visual processing, each ganglion cell combines, analyzes, and encodes the information from the photoreceptors in its receptive field before transmitting the information to the brain (Ringach, 2009).
bipolar cells
In the retina, the specialized neurons that connect the rods and cones with the ganglion cells.
Signals from rods and signals from cones are processed differently in the ganglion. For the most part, a single ganglion cell receives information from only one or two cones but might well receive information from a hundred or more rods. The messages from these many different rods are combined in the retina before they are sent to the brain. Thus, the brain receives less specific visual information from the rods and messages of much greater visual detail from the cones.
As an analogy to how rod information is processed, imagine listening to a hundred people trying to talk at once over the same telephone line. You would hear the sound of many people talking, but individual voices would be blurred. Now imagine listening to the voice of a single individual being transmitted across the same telephone line. Every syllable and sound would be clear and distinct. In much the same way, cones use the ganglion cells to provide the brain with more specific visual information than is received from rods.
Because of this difference in how information is processed, cones are especially important in visual acuity—the ability to see fine details. Visual acuity is strongest when images are focused on the fovea because of the high concentration of cones there.
FROM EYE TO BRAIN
How is information transmitted from the ganglion cells of the retina to the brain? The 1 million axons of the ganglion cells are bundled together to form the optic nerve, a thick nerve that exits from the back of the eye at the optic disk and extends to the brain (see FIGURE 3.5). The optic nerve has about the same diameter as a pencil. After exiting the eyes, the left and right optic nerves meet at the optic chiasm. Then the fibers of the left and right optic nerves split in two. One set of axons crosses over and projects to the opposite side of the brain. The other set of axons forms a pathway that continues along the same side of the brain (see FIGURE 3.5).
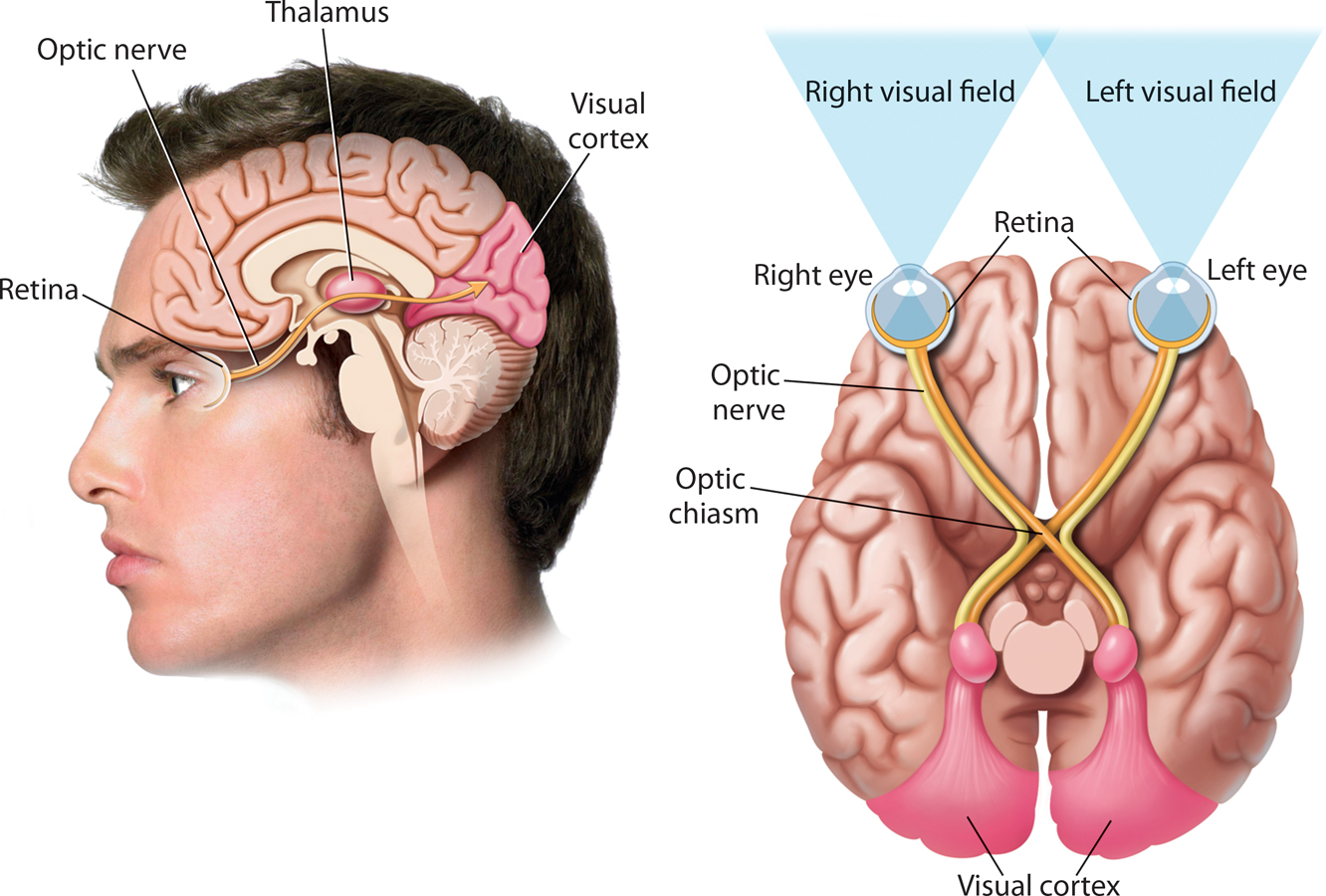
optic nerve
The thick nerve that exits from the back of the eye and carries visual information to the visual cortex in the brain.
optic chiasm
(KY-az-uhm) The point in the brain where the optic nerve fibers from each eye meet and partly cross over to the opposite side of the brain.
From the optic chiasm, most of the optic nerve axons project to the brain structure called the thalamus. (For more on the specific brain structures involved in vision, see Chapter 2). This primary pathway seems to be responsible for processing information about form, color, brightness, and depth. A smaller number of axons follow a detour to areas in the midbrain before they make their way to the thalamus. This secondary pathway seems to be involved in processing information about the location of an object.
Neuroscientists now know that there are several distinct neural pathways in the visual system, each responsible for handling a different aspect of vision (Paik & Ringach, 2011; Purves, 2009). Although specialized, the separate pathways are highly interconnected. From the thalamus, the signals are sent to the visual cortex, where they are decoded and interpreted.
Most of the receiving neurons in the visual cortex of the brain are highly specialized. Each responds to a particular type of visual stimulation, such as angles, edges, lines, and other forms, and even to the movement and distance of objects (Hubel & Wiesel, 2005; Livingstone & Hubel, 1988). These neurons are sometimes called feature detectors because they detect, or respond to, particular features or aspects of more complex visual stimuli. Reassembling the features into a recognizable image involves additional levels of processing in the visual cortex and other regions of the brain, including the frontal lobes.
Understanding exactly how neural responses of individual feature detection cells become integrated into the visual perceptions of faces and objects is a major goal of contemporary neuroscience (Celesia, 2010; Mahon & Caramazza, 2011). Experience also plays an important role in the development of perception. In the Focus on Neuroscience, we explore how Mike May’s perceptual abilities were affected by his lack of visual experience.
Color Vision
We see images of an apple, a banana, and an orange because these objects reflect light waves. But why do we perceive that the apple is red and the banana yellow? What makes an orange orange?
FOCUS ON NEUROSCIENCE
Vision, Experience, and the Brain
After Mike’s surgery, his retina and optic nerve were completely normal. Formal testing showed that Mike had excellent color perception and that he could easily identify simple shapes and lines that were oriented in different directions. These abilities correspond to visual pathways that develop very early. Mike’s motion perception was also very good. When thrown a ball, he could catch it more than 80 percent of the time.
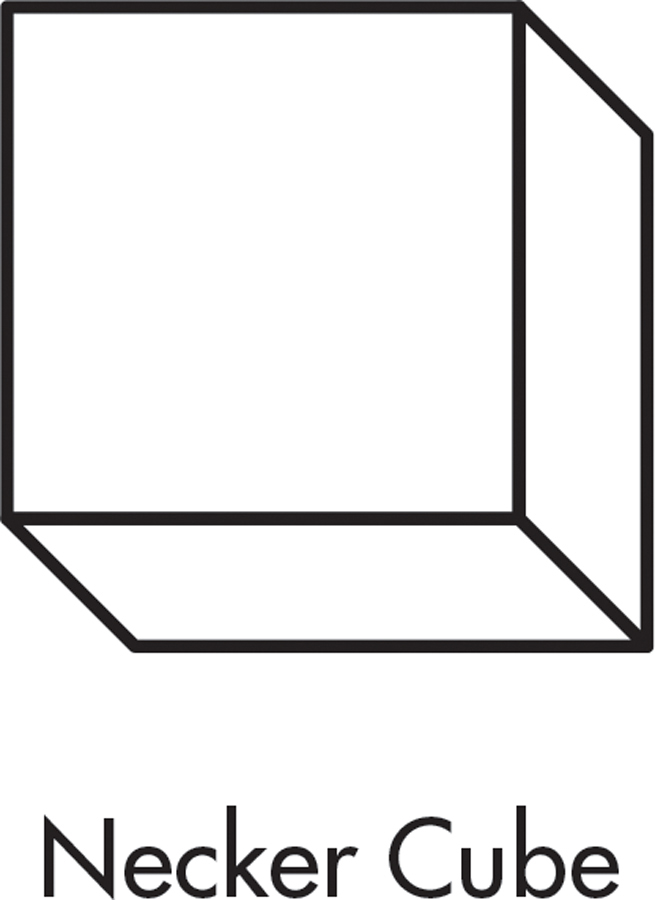
Perceiving and identifying common objects, however, was difficult. Although Mike could “see” an object, he had to consciously use visual cues to work out its identity. For example, when shown the simple drawing above right, called a “Necker cube,” Mike described it as “a square with lines.” But when shown the same image as a rotating image on a computer screen, Mike immediately identified it as a cube. Functional MRI scans showed that Mike’s brain activity was nearly normal when shown a moving object.
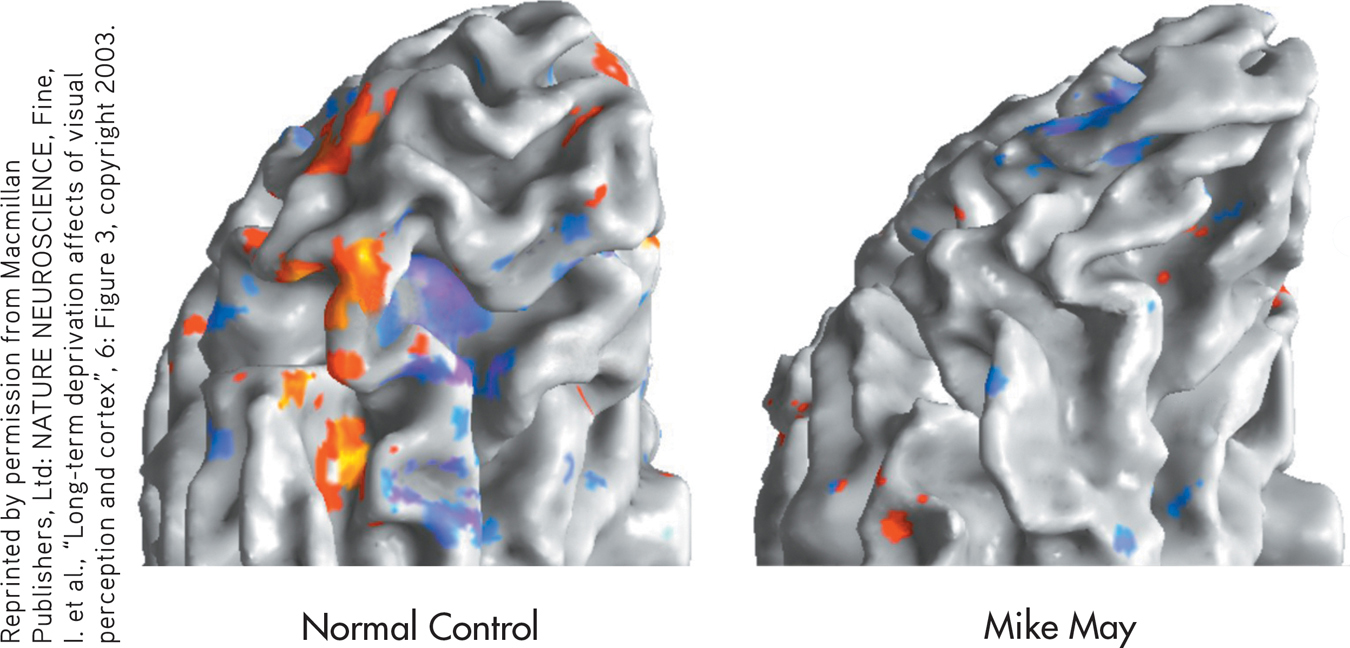
What about more complex objects, like faces? Even three years after his surgery, Mike recognizes his wife and sons by their hair color, gait, and other clues, not by their faces. He can’t tell whether a face is male or female, or whether its expression is happy or sad. Functional MRI scans revealed that when Mike is shown faces or objects, the part of the brain that is normally activated is silent (see brain scans).
For people with normal vision, recognizing complex three-dimensional objects—like tables, shoes, trees, or pencils—is automatic. But as Mike’s story shows, these perceptual conclusions are actually based on experience and built up over time.
Neuroscientist Ione Fine and her colleagues (2003, 2008), who have studied Mike’s visual abilities, believe that Mike’s case indicates that some visual pathways develop earlier than others. Color and motion perception, they point out, develop early in infancy. But because people will continue to encounter new objects and faces throughout life, areas of the brain that are specialized to process faces and objects show plasticity. In Mike’s case, these brain centers never developed.
MYTH  SCIENCE
SCIENCE
Is it true that an object’s color is not an intrinsic property of the object?
THE EXPERIENCE OF COLORWHAT MAKES AN ORANGE ORANGE?
Color is not a property of an object, but a sensation perceived in the brain (Werner & others, 2007). To explain how we perceive color, we must return to the original visual stimulus—light.
Our experience of color involves three properties of the light wave. First, what we usually refer to as color is a property more accurately termed hue. Hue varies with the wavelength of light. Look again at FIGURE 3.2 on page 90. Different wavelengths correspond to our subjective experience of different colors. Wavelengths of about 400 nanometers are perceived as violet. Wavelengths of about 700 nanometers are perceived as red. In between are orange, yellow, green, blue, and indigo.
color
The perceptual experience of different wavelengths of light, involving hue, saturation (purity), and brightness (intensity).
hue
The property of wavelengths of light known as color; different wavelengths correspond to our subjective experience of different colors.
Second, the saturation, or purity, of the color corresponds to the purity of the light wave. Pure red, for example, produced by a single wavelength, is more saturated than pink, which is produced by a combination of wavelengths (red plus white light). In everyday language, saturation refers to the richness of a color. A highly saturated color is vivid and rich; a less saturated color is faded and washed out.
saturation
The property of color that corresponds to the purity of the light wave.
The third property of color is brightness, or perceived intensity. Brightness corresponds to the amplitude of the light wave: The higher the amplitude, the greater the degree of brightness.
brightness
The perceived intensity of a color, which corresponds to the amplitude of the light wave.
These three properties of color—hue, saturation, and brightness—are responsible for the amazing range of colors we experience. A person with normal color vision can discriminate from 120 to 150 color differences based on differences in hue, or wavelength, alone. When saturation and brightness are also factored in, we can potentially perceive millions of different colors (Elliot & Maier, 2014).
Many people mistakenly believe that white light contains no color. White light actually contains all wavelengths—and thus all colors—of the visible part of the electromagnetic spectrum. A glass prism placed in sunlight creates a rainbow because it separates sunlight into all the colors of the visible light spectrum.
So we’re back to the question: What makes an orange orange? Intuitively, it seems obvious that the color of any object is an inseparable property of the object—unless we spill paint or spaghetti sauce on it. In reality, color is a sensation perceived in the brain (Werner & others, 2007).
Our perception of color is primarily determined by the wavelength of light that an object reflects. If your T-shirt is red, it’s red because the cloth is reflecting only the wavelength of light that corresponds to the red portion of the spectrum. The T-shirt is absorbing the wavelengths that correspond to all other colors. An object appears white because it reflects all the wavelengths of visible light and absorbs none. An object appears black when it absorbs all the wavelengths of visible light and reflects none. Of course, in everyday life, our perceptions of color are also strongly affected by the amount or type of light falling on an object or the textures and colors that surround it (Purves, 2009; Shevell & Kingdom, 2008).

HOW WE SEE COLOR
Color vision has interested scientists for hundreds of years. The first scientific theory of color vision, proposed by Hermann von Helmholtz (1821–
The Trichromatic TheoryAs you’ll recall, only the cones are involved in color vision. According to the trichromatic theory of color vision, there are three varieties of cones. Each type of cone is especially sensitive to certain wavelengths—red light (long wavelengths), green light (medium wavelengths), or blue light (short wavelengths). For the sake of simplicity, we will refer to red-sensitive, green-sensitive, and blue-sensitive cones, but keep in mind that there is some overlap in the wavelengths to which a cone is sensitive (Purves, 2009). A given cone will be very sensitive to one of the three colors and only slightly responsive to the other two.
trichromatic theory of color vision
The theory that the sensation of color results because cones in the retina are especially sensitive to red light (long wavelengths), green light (medium wavelengths), or blue light (short wavelengths).
When a color other than red, green, or blue strikes the retina, it stimulates a combination of cones. For example, if yellow light strikes the retina, both the red-sensitive and green-sensitive cones are stimulated; purple light evokes strong reactions from red-sensitive and blue-sensitive cones. The trichromatic theory of color vision received compelling research support in 1964, when George Wald showed that different cones were indeed activated by red, blue, and green light.
The trichromatic theory provides a good explanation for the most common form of color blindness: red–
color blindness
One of several inherited forms of color deficiency or weakness in which an individual cannot distinguish between certain colors.
The Opponent-Process Theory

The trichromatic theory cannot account for all aspects of color vision. One important phenomenon that the theory does not explain is the afterimage. An afterimage is a visual experience that occurs after the original source of stimulation is no longer present. To experience an afterimage firsthand, follow the instructions in FIGURE 3.6. What do you see?
afterimage
A visual experience that occurs after the original source of stimulation is no longer present.
Afterimages can be explained by the opponent-process theory of color vision, which proposes a different mechanism of color detection from the one set forth in the trichromatic theory. According to the opponent-process theory of color vision, there are four basic colors, which are divided into two pairs of color-sensitive neurons: red–
opponent-process theory of color vision
The theory that color vision is the product of opposing pairs of color receptors: red–
For example, red light evokes a response of RED-YES–
Afterimages can be explained when the opponent-process theory is combined with the general principle of sensory adaptation (Jameson & Hurvich, 1989). If you stare continuously at one color, sensory adaptation eventually occurs and your visual receptors become less sensitive to that color. What happens when you subsequently stare at a white surface?
If you remember that white light is made up of the wavelengths for all colors, you may be able to predict the result. The receptors for the original color have adapted to the constant stimulation and are temporarily “off duty.” Thus, they do not respond to that color. Instead, only the receptors for the opposing color will be activated, and you perceive the wavelength of only the opposing color. For example, if you stare at a patch of green, your green receptors eventually become “tired.” The wavelengths for both green and red light are reflected by the white surface, but since the green receptors are “off,” only the red receptors are activated. Staring at the green, black, and yellow flag in FIGURE 3.6 should have produced an afterimage of opposing colors: a red, white, and blue American flag.

An Integrated Explanation of Color VisionAt the beginning of this section, we said that current research has shown that both the trichromatic theory and the opponent-process theory of color vision are accurate. How can both theories be right? It turns out that each theory correctly describes color vision at a different level of visual processing.
As described by the trichromatic theory, the cones of the retina do indeed respond to and encode color in terms of red, green, and blue. But recall that signals from the cones and rods are partially processed in the ganglion cells before being transmitted along the optic nerve to the brain. Researchers now believe that an additional level of color processing takes place in the ganglion cells (Demb & Brainard, 2010).
As described by the opponent-process theory, the ganglion cells respond to and encode color in terms of opposing pairs (DeValois & DeValois, 1975; Solomon & Lennie, 2007). In the brain, the thalamus and visual cortex also encode color in terms of opponent pairs. Consequently, both theories contribute to our understanding of the process of color vision. Each theory simply describes color vision at a different stage of visual processing (Hubel, 1995; Werner & others, 2007).
CONCEPT REVIEW 3.1
Principles of Sensation and Vision
Fill in the missing words to complete each statement.
Question 3.1
| 1. | Sensory receptors convert different forms of physical energy into neural signals through the process of . |
Question 3.2
| 2. | The sensory receptors for vision are contained in the . |
Question 3.3
| 3. | Complete the sequence of visual information transmission: rods and cones → cells → cells → optic nerve → → thalamus → . |
Question 3.4
| 4. | According to the
theory of color vision, the person with red– |
Question 3.5
| 5. | When people stare at a yellow circle and then shift their eyes to a white surface, the afterimage of the circle appears . This phenomenon can best be explained by the theory of color vision. |
Test your understanding of Sensation vs. Perception; Vision with
 .
.