SUMMARY
Genetic research and plant and animal breeding often necessitate the synthesis of genotypes that are complex combinations of alleles from different genes. Such genes can be on the same chromosome or on different chromosomes; the latter is the main subject of this chapter.
In the simplest case—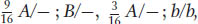
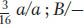 , and
, and 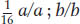 . The 1:1:1:1 and 9:3:3:1 ratios are both diagnostic of independent assortment.
. The 1:1:1:1 and 9:3:3:1 ratios are both diagnostic of independent assortment.
More complex genotypes composed of independently assorting genes can be treated as extensions of the case for single-
Sequential generations of selfing increase the proportions of homozygotes, according to the principles of equal segregation and independent assortment (if the genes are on different chromosomes). Hence, selfing is used to create complex pure lines with combinations of desirable mutations.
The independent assortment of chromosomes at meiosis can be observed cytologically by using heteromorphic chromosome pairs (those that show a structural difference). The X and Y chromosomes are one such case, but other, rarer cases can be found and used for this demonstration. The independent assortment of genes at the level of single meiocytes can be observed in the ascomycete fungi, because the asci show the two alternative types of segregations at equal frequencies.
One of the main functions of meiosis is to produce recom-
Metric characters such as color intensity show a continuous distribution in a population. Continuous distributions can be based on environmental variation or on variant alleles of multiple genes or on a combination of both. A simple genetic model proposes that the active alleles of several genes (called polygenes) contribute more or less additively to the metric character. In an analysis of the progeny from the self of a multiply heterozygous individual, the histogram showing the proportion of each phenotype approximates a bell-
The small subsets of the genome found in mitochondria and chloroplasts are inherited independently of the nuclear genome. Mutants in these organelle genes often show maternal inheritance, along with the cytoplasm, which is the location of these organelles. In genetically mixed cytoplasms (cytohets), the two genotypes (say, wild type and mutant) often sort themselves out into different daughter cells by a poorly understood process called cytoplasmic segregation. Mitochondrial mutation in humans results in diseases that show cytoplasmic segregation in body tissues and maternal inheritance in a mating.