Chapter Introduction
Mapping Eukaryote Chromosomes by Recombination
127
Mapping Eukaryote
Chromosomes by
Recombination
CHAPTER
4
LEARNING OUTCOMES
After completing this chapter, you will be able to
Perform a quantitative analysis of the progeny of a dihybrid testcross to assess whether or not the two genes are linked on the same chromosome.
Extend the same type of analysis to several loci to produce a map of the relative positions of loci on a chromosome.
In ascomycete fungi, map the centromeres to other linked loci.
In asci, predict allele ratios stemming from specific steps in the heteroduplex model of crossing over.
OUTLINE
|
4.1 |
Diagnostics of linkage |
|
4.2 |
Mapping by recombinant frequency |
|
4.3 |
Mapping with molecular markers |
|
4.4 |
Centromere mapping with linear tetrads |
|
4.5 |
Using the chi-square test to infer linkage |
|
4.6 |
Accounting for unseen multiple crossovers |
|
4.7 |
Using recombination-based maps in conjunction with physical maps |
|
4.8 |
The molecular mechanism of crossing over |
128
Some of the questions that geneticists want to answer about the genome are, What genes are present in the genome? What functions do they have? What positions do they occupy on the chromosomes? Their pursuit of the third question is broadly called mapping. Mapping is the main focus of this chapter, but all three questions are interrelated, as we will see.
We all have an everyday feeling for the importance of maps in general, and, indeed, we have all used them at some time in our lives to find our way around. Relevant to the focus of this chapter is that, in some situations, several maps need to be used simultaneously. A good example in everyday life is in navigating the dense array of streets and buildings of a city such as London, England. A street map that shows the general layout is one necessity. However, the street map is used by tourists and Londoners alike in conjunction with another map, that of the underground railway system. The underground system is so complex and spaghetti-
129
Obtaining a map of gene positions on the chromosomes is an endeavor that has occupied thousands of geneticists for the past 80 years or so. Why is it so important? There are several reasons:
Gene position is crucial information needed to build complex genotypes required for experimental purposes or for commercial applications. For example, in Chapter 6, we will see cases in which special allelic combinations must be put together to explore gene interaction.
Knowing the position occupied by a gene provides a way of discovering its structure and function. A gene’s position can be used to define it at the DNA level. In turn, the DNA sequence of a wild-
type gene or its mutant allele is a necessary part of deducing its underlying function. The genes present and their arrangement on chromosomes are often slightly different in related species. For example, the rather long human chromosome number 2 is split into two shorter chromosomes in the great apes. By comparing such differences, geneticists can deduce the evolutionary genetic mechanisms through which these genomes diverged. Hence, chromosome maps are useful in interpreting mechanisms of evolution.
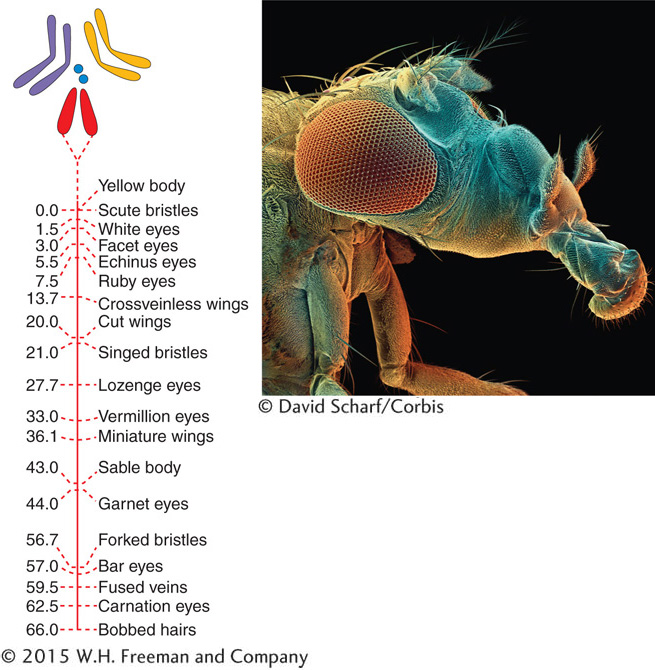
The arrangement of genes on chromosomes is represented diagrammatically as a unidimensional chromosome map, showing gene positions, known as loci (sing., locus), and the distances between the loci based on some kind of scale. Two basic types of chromosome maps are currently used in genetics; they are assembled by quite different procedures yet are used in a complementary way. Recombination-